

Engineered Tissue Folding by Mechanical Compaction of the Mesenchyme
- PMID: 29290586
- PMCID: PMC5826757
- DOI: 10.1016/j.devcel.2017.12.004
Engineered Tissue Folding by Mechanical Compaction of the Mesenchyme
Abstract
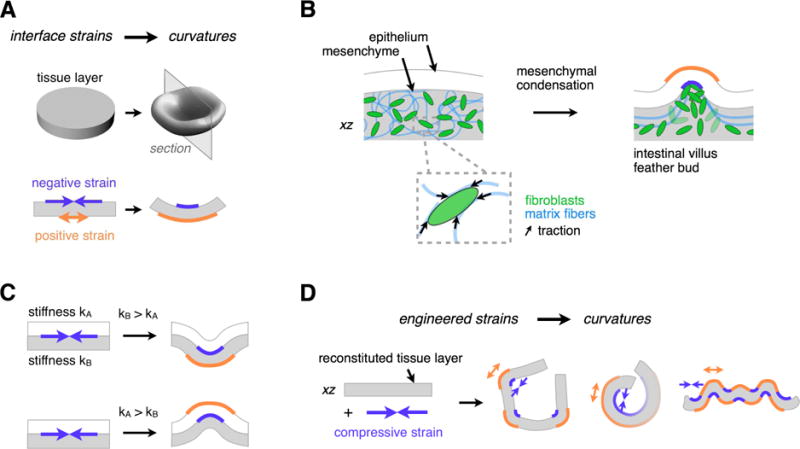
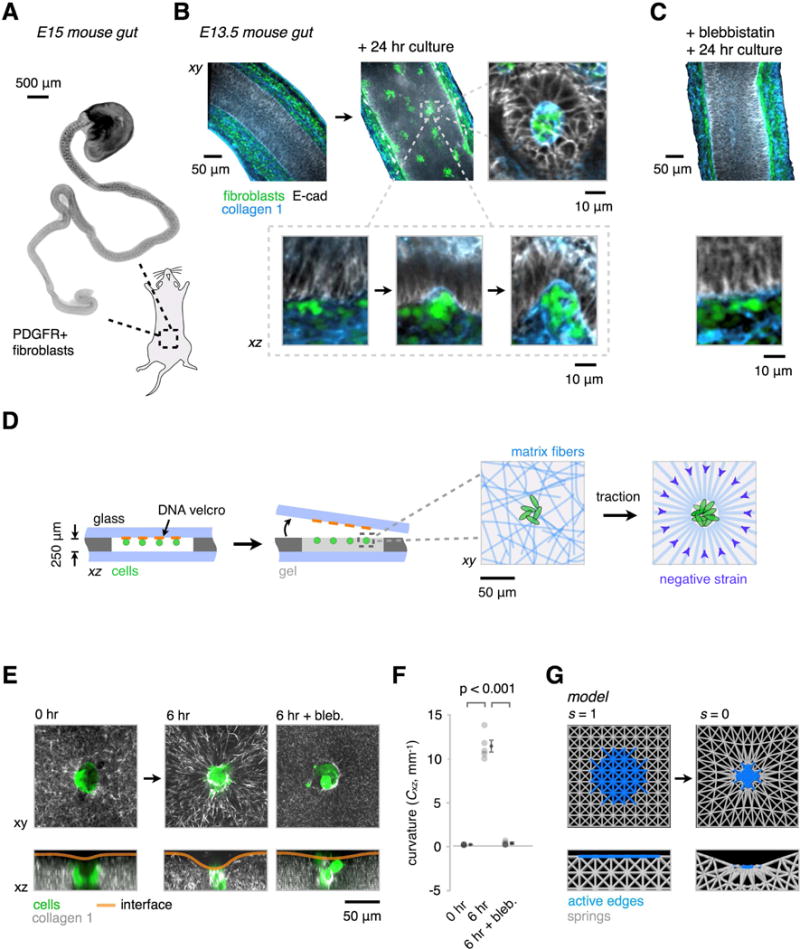
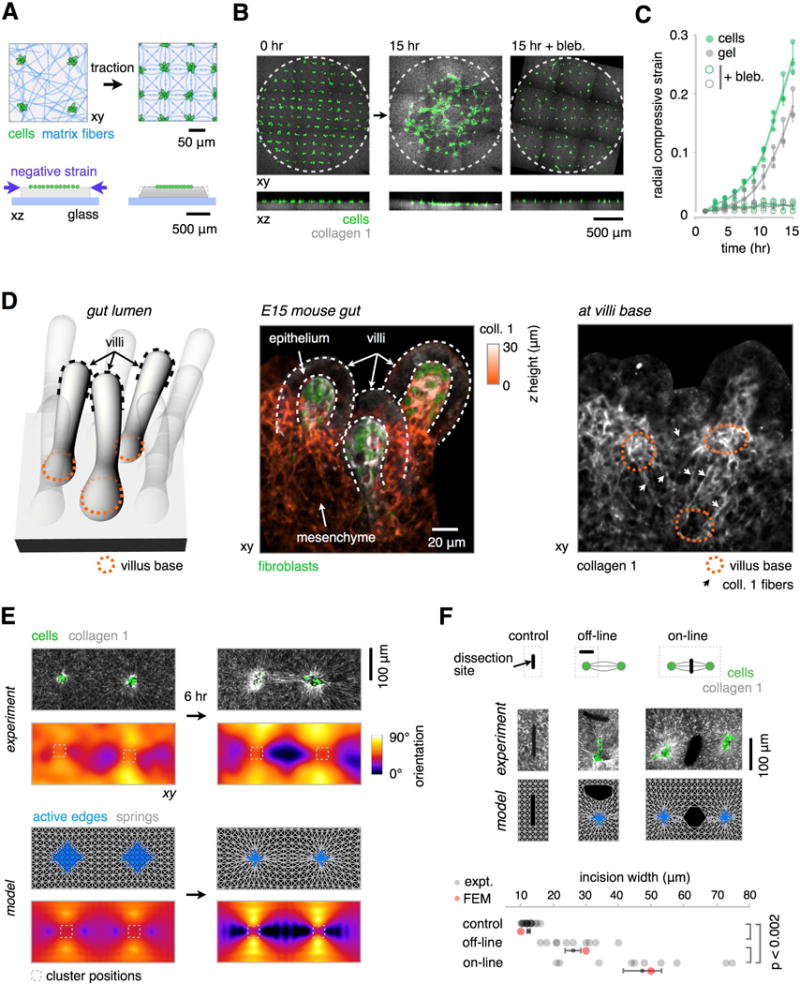
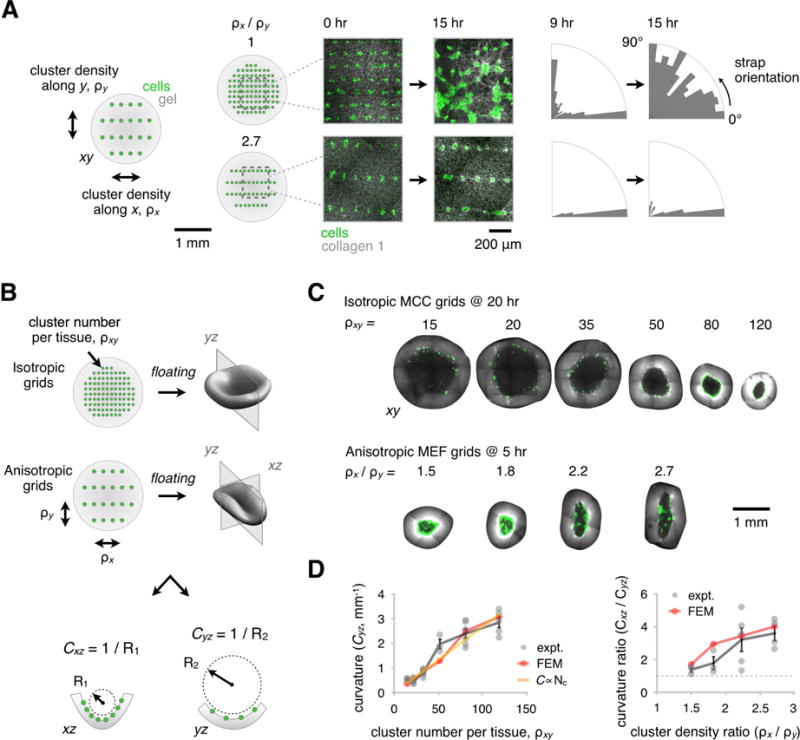
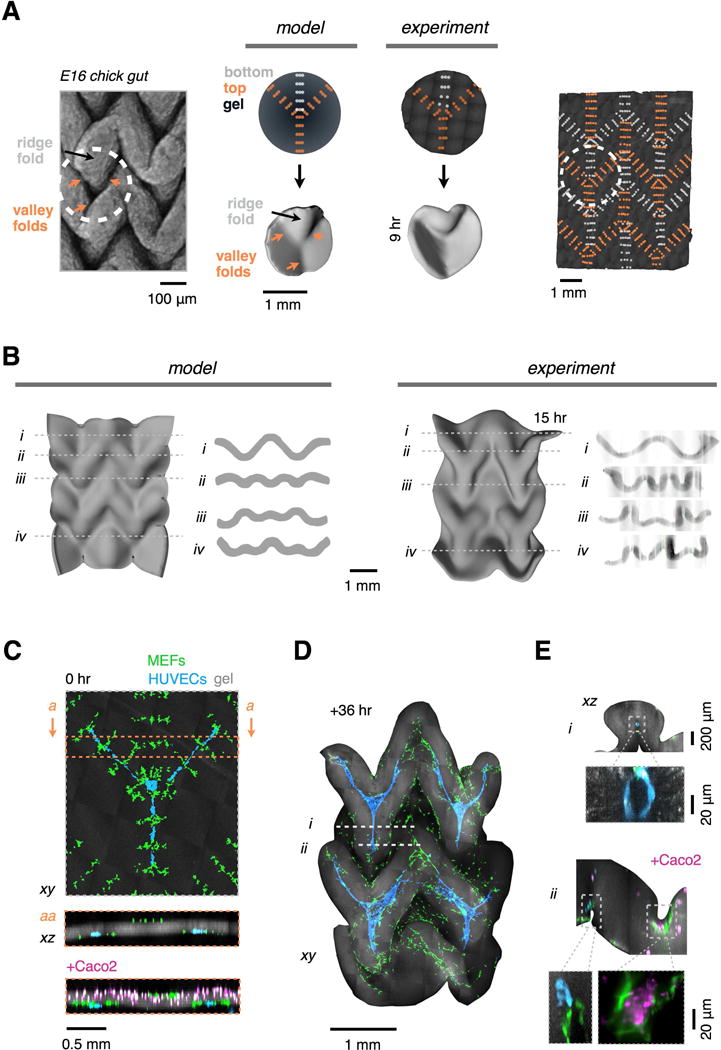
Many tissues fold into complex shapes during development. Controlling this process in vitro would represent an important advance for tissue engineering. We use embryonic tissue explants, finite element modeling, and 3D cell-patterning techniques to show that mechanical compaction of the extracellular matrix during mesenchymal condensation is sufficient to drive tissue folding along programmed trajectories. The process requires cell contractility, generates strains at tissue interfaces, and causes patterns of collagen alignment around and between condensates. Aligned collagen fibers support elevated tensions that promote the folding of interfaces along paths that can be predicted by modeling. We demonstrate the robustness and versatility of this strategy for sculpting tissue interfaces by directing the morphogenesis of a variety of folded tissue forms from patterns of mesenchymal condensates. These studies provide insight into the active mechanical properties of the embryonic mesenchyme and establish engineering strategies for more robustly directing tissue morphogenesis ex vivo.
Keywords: intestinal villi; mechanobiology; mesenchymal condensation; smart materials; soft matter; soft robotics; synthetic biology; tissue engineering; tissue folding.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
A.J.H. and Z.J.G. are inventors on intellectual property related to engineered tissue folding.
Figures

Comment in
-
Engineering Control over 3D Morphogenesis by Tissue Origami.Dev Cell. 2018 Jan 22;44(2):131-132. doi: 10.1016/j.devcel.2018.01.005. Dev Cell. 2018. PMID: 29401414
-
Engineering Biology by Controlling Tissue Folding.Trends Biotechnol. 2018 Apr;36(4):341-343. doi: 10.1016/j.tibtech.2018.02.003. Epub 2018 Feb 22. Trends Biotechnol. 2018. PMID: 29478676
References
-
- Armon S, Efrati E, Kupferman R, Sharon E. Geometry and Mechanics in the Opening of Chiral Seed Pods. Science. 2011;333:1726–1730. - PubMed
-
- Bender J, Müller M, Otaduy MA, Teschner M, Macklin M. A Survey on Position-Based Simulation Methods in Computer Graphics. Computer Graphics Forum. 2014;33:228–251.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
