

The numerical abilities of anurans and their neural correlates: insights from neuroethological studies of acoustic communication
- PMID: 29292359
- PMCID: PMC5784039
- DOI: 10.1098/rstb.2016.0512
The numerical abilities of anurans and their neural correlates: insights from neuroethological studies of acoustic communication
Abstract
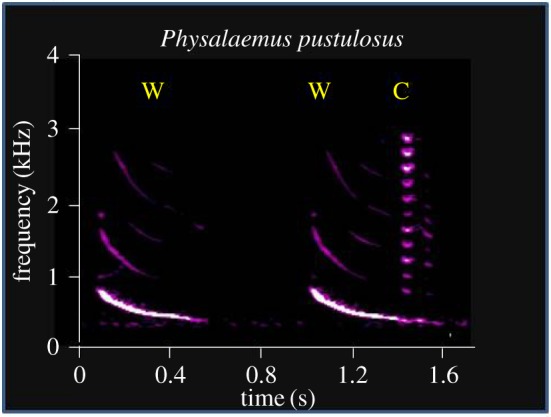
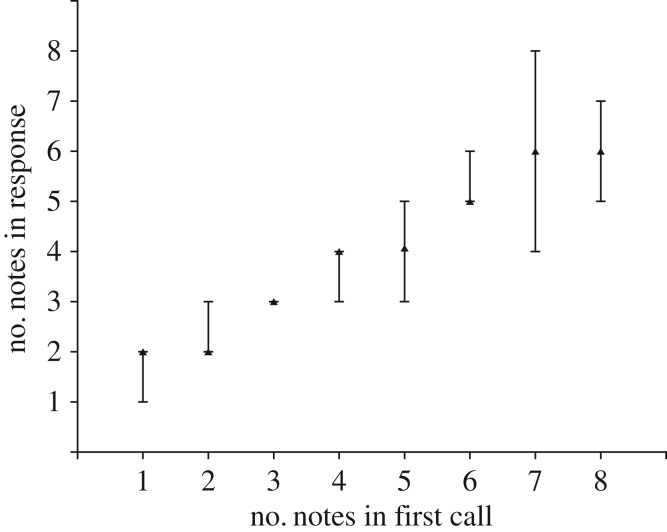
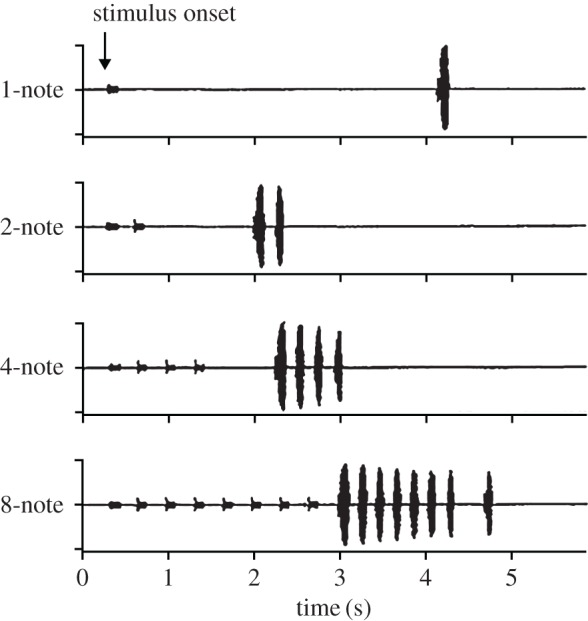
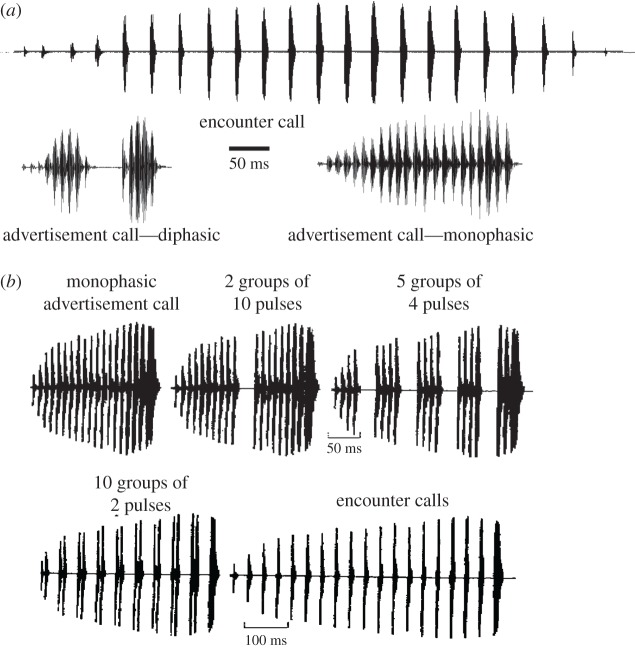
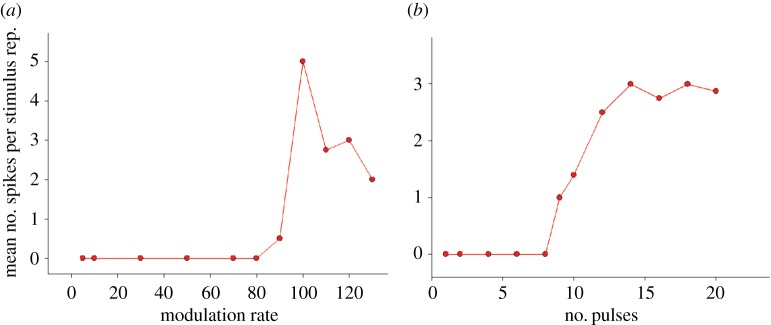
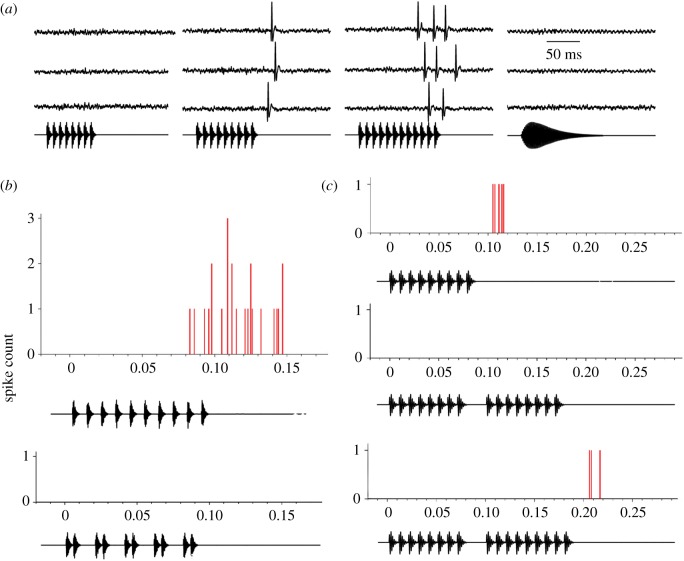
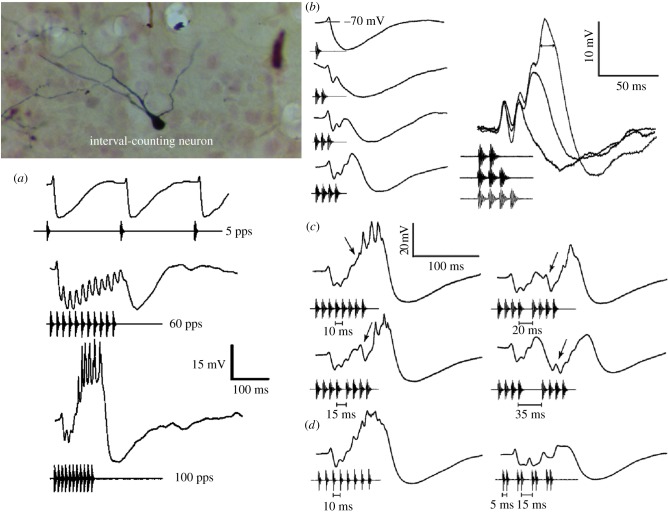
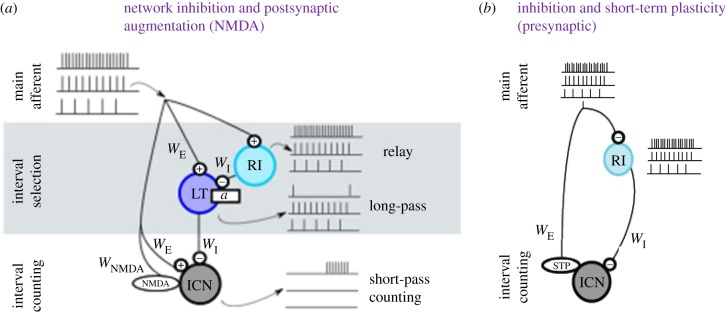
Acoustic communication is important in the reproductive behaviour of anurans. The acoustic repertoire of most species consists of several call types, but some anurans gradually increase the complexity of their calls during aggressive interactions between males and when approached by females. In these interactions, males may closely match the number of calls or notes in a sequence that a neighbour produces, thereby revealing their numerical abilities. Anurans are also able to discern the number of sequential properly timed pulses (notes). The temporal intervals between successive pulses provide information about species identity and call type. A neural correlate of this numerical ability is evident in the responses of 'interval-counting' neurons, which show 'tuning' for intermediate to fast pulse rates and respond only after at least a threshold number of pulses have occurred with the correct timing. A single interpulse interval that is two to three times the optimal value can reset this interval-counting process. Whole-cell recordings from midbrain neurons, in vivo, have revealed that complex interplay between activity-dependent excitation and inhibition contributes to this counting process. Single pulses primarily elicit inhibition. As additional pulses are presented with optimal intervals, cells become progressively depolarized and spike after a threshold number of intervals have occurred.This article is part of a discussion meeting issue 'The origins of numerical abilities'.
Keywords: acoustic communication; auditory; call matching; frogs; inferior colliculus; interval-counting neurons.
© 2017 The Author(s).
Conflict of interest statement
I declare I have no competing interests.
Figures
References
-
- Wells KD, Schwartz JJ. 2006. The behavioral ecology of anuran communication. In Hearing and sound communication in amphibians (eds Narins PM, Feng AS, Fay R, Popper A), pp. 44–86. Berlin, Germany: Springer.
-
- Wells KD. 1977. The social behaviour of anuran amphibians. Anim. Behav. 25, 666–693. (10.1016/0003-3472(77)90118-X) - DOI
-
- Rose GJ, Brenowitz EA. 1997. Plasticity of aggressive thresholds in Hyla regilladiscrete accommodation to encounter calls. Anim. Behav. 53, 353–361. (10.1006/anbe.1996.0400) - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
