

A role for small RNA in regulating innate immunity during plant growth
- PMID: 29293695
- PMCID: PMC5766230
- DOI: 10.1371/journal.ppat.1006756
A role for small RNA in regulating innate immunity during plant growth
Abstract
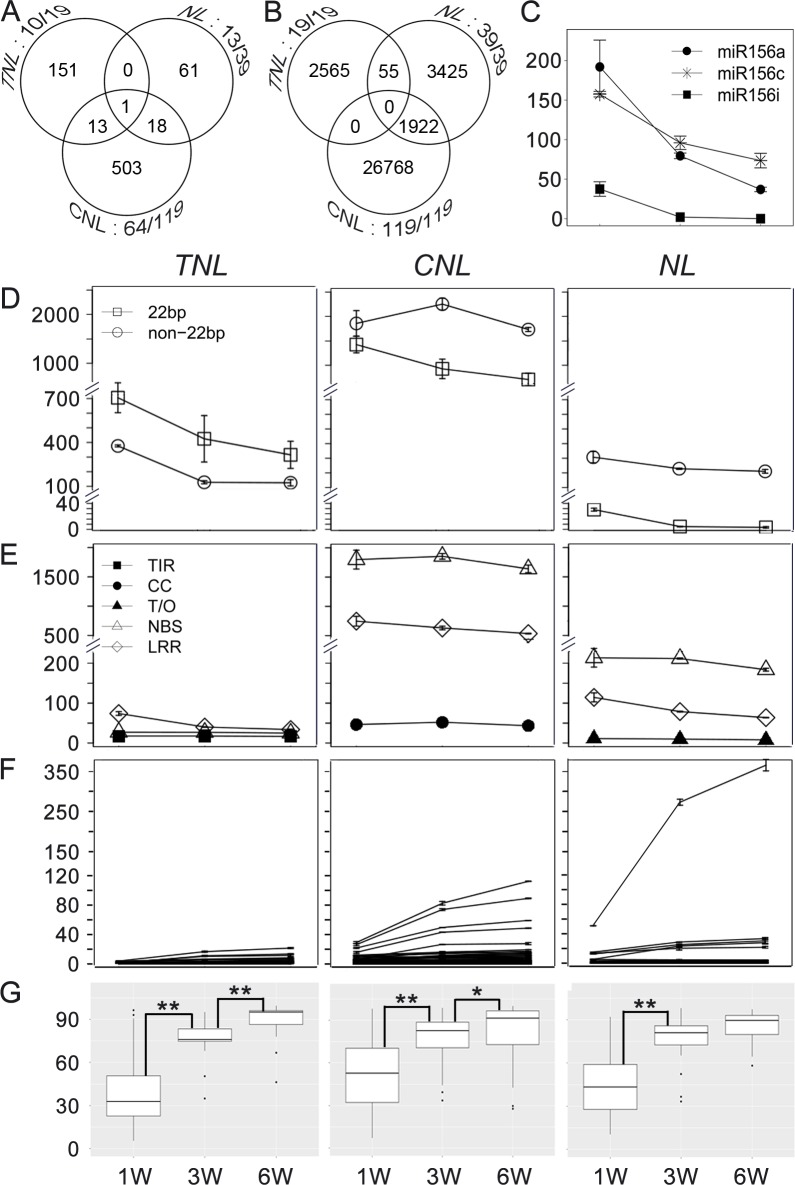
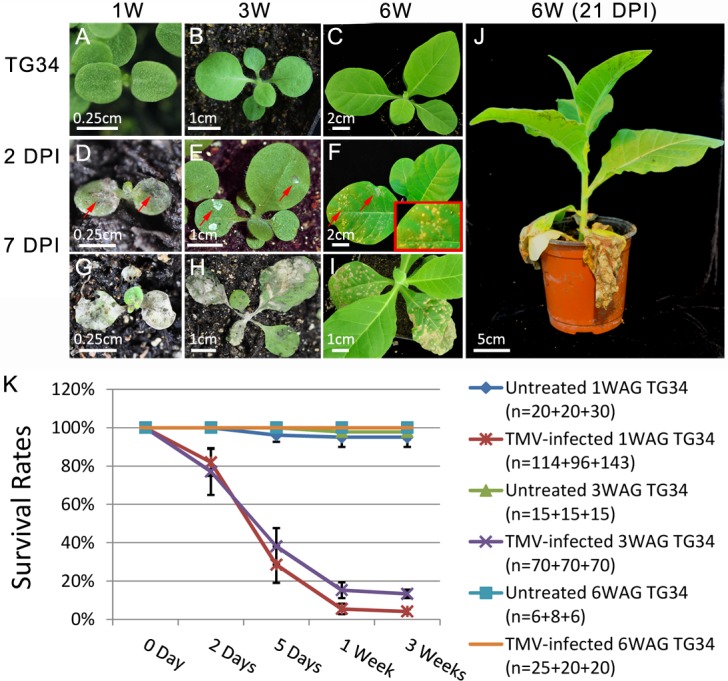
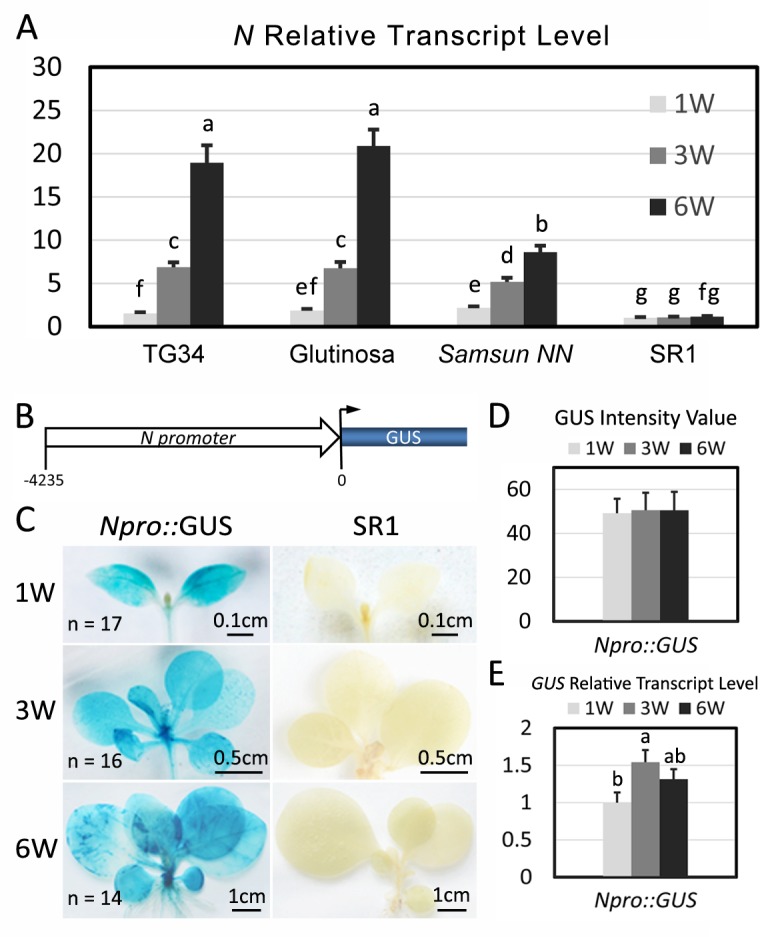
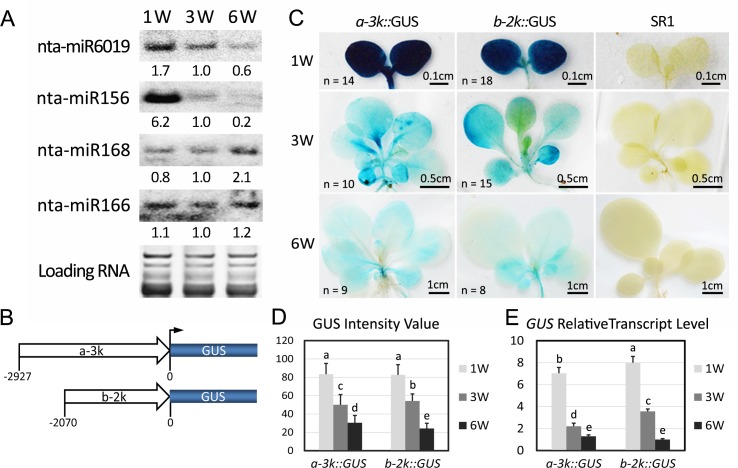
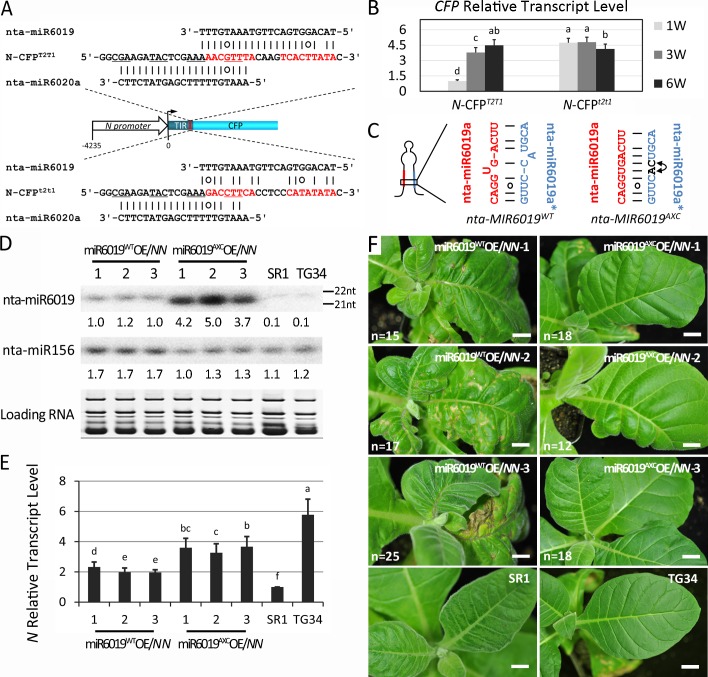
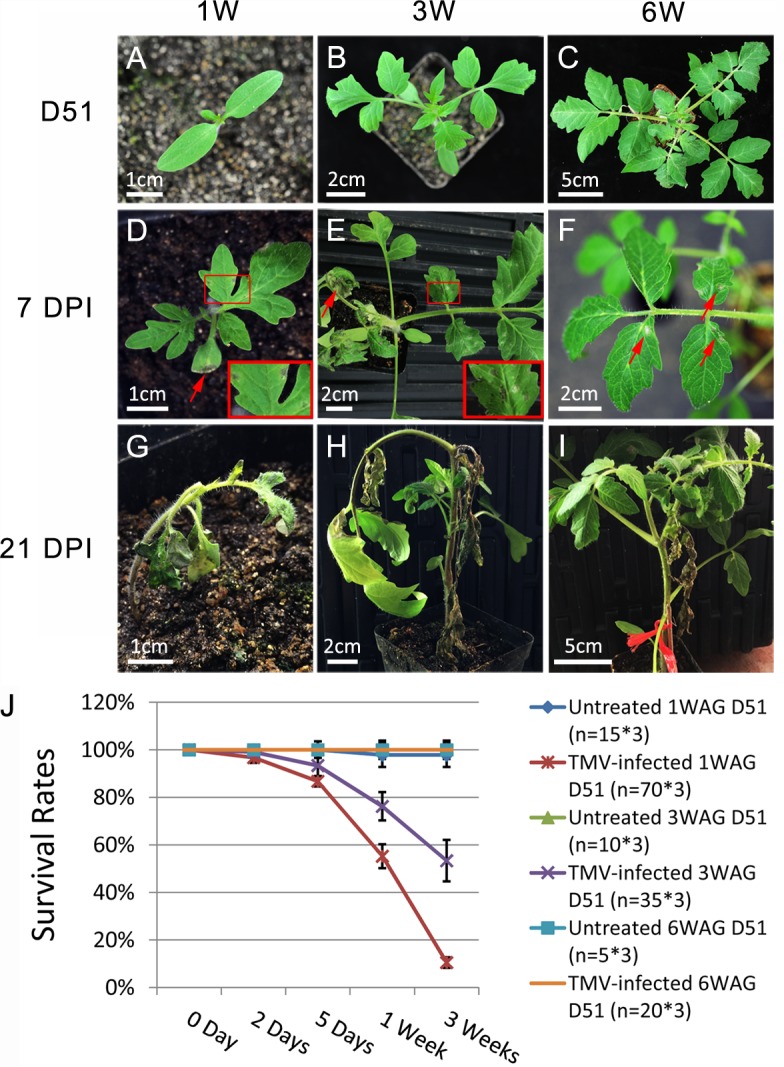
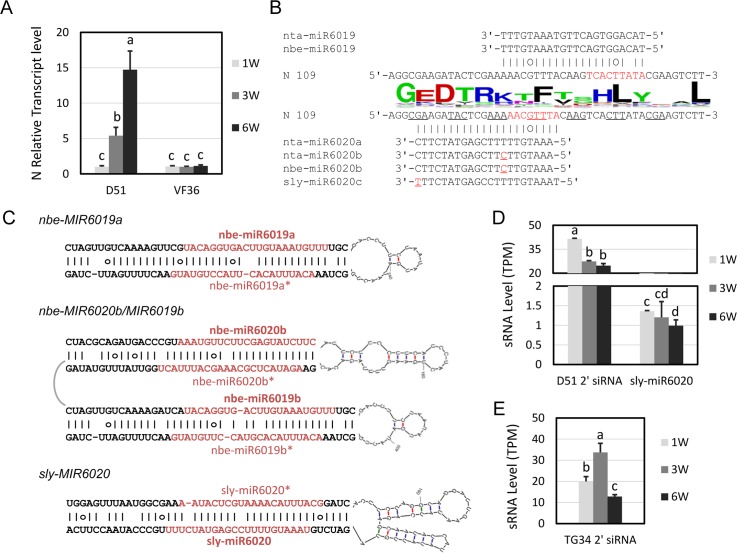
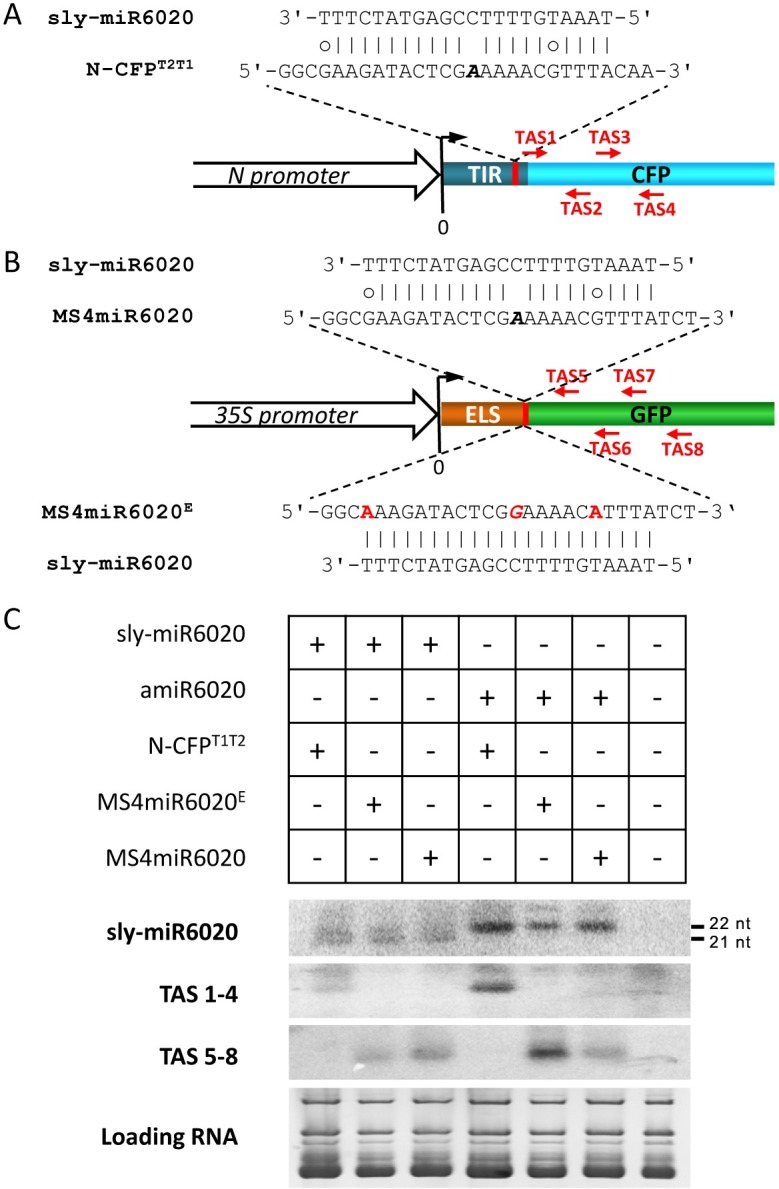
Plant genomes encode large numbers of nucleotide-binding (NB) leucine-rich repeat (LRR) immune receptors (NLR) that mediate effector triggered immunity (ETI) and play key roles in protecting crops from diseases caused by devastating pathogens. Fitness costs are associated with plant NLR genes and regulation of NLR genes by micro(mi)RNAs and phased small interfering RNAs (phasiRNA) is proposed as a mechanism for reducing these fitness costs. However, whether NLR expression and NLR-mediated immunity are regulated during plant growth is unclear. We conducted genome-wide transcriptome analysis and showed that NLR expression gradually increased while expression of their regulatory small RNAs (sRNA) gradually decreased as plants matured, indicating that sRNAs could play a role in regulating NLR expression during plant growth. We further tested the role of miRNA in the growth regulation of NLRs using the tobacco mosaic virus (TMV) resistance gene N, which was targeted by miR6019 and miR6020. We showed that N-mediated resistance to TMV effectively restricted this virus to the infected leaves of 6-week old plants, whereas TMV infection was lethal in 1- and 3-week old seedlings due to virus-induced systemic necrosis. We further found that N transcript levels gradually increased while miR6019 levels gradually decreased during seedling maturation that occurs in the weeks after germination. Analyses of reporter genes in transgenic plants showed that growth regulation of N expression was post-transcriptionally mediated by MIR6019/6020 whereas MIR6019/6020 was regulated at the transcriptional level during plant growth. TMV infection of MIR6019/6020 transgenic plants indicated a key role for miR6019-triggered phasiRNA production for regulation of N-mediated immunity. Together our results demonstrate a mechanistic role for miRNAs in regulating innate immunity during plant growth.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
MicroRNA regulation of plant innate immune receptors.Proc Natl Acad Sci U S A. 2012 Jan 31;109(5):1790-5. doi: 10.1073/pnas.1118282109. Epub 2012 Jan 18. Proc Natl Acad Sci U S A. 2012. PMID: 22307647 Free PMC article.
-
Mitochondrial alternative oxidase is involved in both compatible and incompatible host-virus combinations in Nicotiana benthamiana.Plant Sci. 2015 Oct;239:26-35. doi: 10.1016/j.plantsci.2015.07.009. Epub 2015 Jul 19. Plant Sci. 2015. PMID: 26398788
-
HC-Pro silencing suppressor significantly alters the gene expression profile in tobacco leaves and flowers.BMC Plant Biol. 2011 Apr 20;11:68. doi: 10.1186/1471-2229-11-68. BMC Plant Biol. 2011. PMID: 21507209 Free PMC article.
-
Animal NLRs provide structural insights into plant NLR function.Ann Bot. 2017 Mar 1;119(5):827-702. doi: 10.1093/aob/mcw171. Ann Bot. 2017. PMID: 27562749 Free PMC article. Review.
-
Fine-Tuning Immunity: Players and Regulators for Plant NLRs.Trends Plant Sci. 2020 Jul;25(7):695-713. doi: 10.1016/j.tplants.2020.02.008. Epub 2020 Mar 17. Trends Plant Sci. 2020. PMID: 32526174 Review.
Cited by
-
Advances in understanding the interaction between Solanaceae NLR resistance proteins and the viral effector Avr.Plant Signal Behav. 2024 Dec 31;19(1):2382497. doi: 10.1080/15592324.2024.2382497. Epub 2024 Sep 23. Plant Signal Behav. 2024. PMID: 39312190 Free PMC article. Review.
-
Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects.Int J Mol Sci. 2022 Dec 19;23(24):16200. doi: 10.3390/ijms232416200. Int J Mol Sci. 2022. PMID: 36555841 Free PMC article. Review.
-
Cross-Kingdom RNAi of Pathogen Effectors Leads to Quantitative Adult Plant Resistance in Wheat.Front Plant Sci. 2020 Mar 10;11:253. doi: 10.3389/fpls.2020.00253. eCollection 2020. Front Plant Sci. 2020. PMID: 32211008 Free PMC article.
-
Overexpression of Nta-miR6155 confers resistance to Phytophthora nicotianae and regulates growth in tobacco (Nicotiana tabacum L.).Front Plant Sci. 2023 Nov 20;14:1281373. doi: 10.3389/fpls.2023.1281373. eCollection 2023. Front Plant Sci. 2023. PMID: 38053762 Free PMC article.
-
Argonaute4 Modulates Resistance to Fusarium brachygibbosum Infection by Regulating Jasmonic Acid Signaling.Plant Physiol. 2020 Oct;184(2):1128-1152. doi: 10.1104/pp.20.00171. Epub 2020 Jul 28. Plant Physiol. 2020. PMID: 32723807 Free PMC article.
References
-
- Caplan J, Padmanabhan M, Dinesh-Kumar SP. Plant NB-LRR immune receptors: from recognition to transcriptional reprogramming. Cell Host Microbe. 2008;3(3):126–35. doi: 10.1016/j.chom.2008.02.010 . - DOI - PubMed
-
- Meyers BC, Kozik A, Griego A, Kuang H, Michelmore RW. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell. 2003;15(4):809–34. doi: 10.1105/tpc.009308 ; PubMed Central PMCID: PMCPMC152331. - DOI - PMC - PubMed
-
- Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int Immunol. 2009;21(4):317–37. doi: 10.1093/intimm/dxp017 ; PubMed Central PMCID: PMCPMC2721684. - DOI - PMC - PubMed
-
- Kollmann TR, Levy O, Montgomery RR, Goriely S. Innate immune function by Toll-like receptors: distinct responses in newborns and the elderly. Immunity. 2012;37(5):771–83. doi: 10.1016/j.immuni.2012.10.014 ; PubMed Central PMCID: PMCPMC3538030. - DOI - PMC - PubMed
-
- Chen XM. microRNA biogenesis and function in plants. Febs Letters. 2005;579(26):5923–31. doi: 10.1016/j.febslet.2005.07.071 WOS:000233002500013. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
