

On the origins and domestication of the olive: a review and perspectives
- PMID: 29293871
- PMCID: PMC5838823
- DOI: 10.1093/aob/mcx145
On the origins and domestication of the olive: a review and perspectives
Erratum in
-
On the origins and domestication of the olive: a review and perspectives.Ann Bot. 2018 Mar 5;121(3):587-588. doi: 10.1093/aob/mcy002. Ann Bot. 2018. PMID: 29390111 Free PMC article. No abstract available.
Abstract
Background: Unravelling domestication processes is crucial for understanding how species respond to anthropogenic pressures, forecasting crop responses to future global changes and improving breeding programmes. Domestication processes for clonally propagated perennials differ markedly from those for seed-propagated annual crops, mostly due to long generation times, clonal propagation and recurrent admixture with local forms, leading to a limited number of generations of selection from wild ancestors. However, additional case studies are required to document this process more fully.
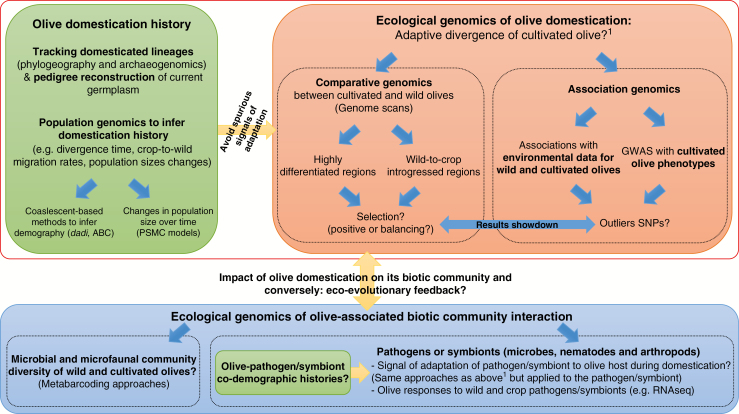
Scope: The olive is an iconic species in Mediterranean cultural history. Its multiple uses and omnipresence in traditional agrosystems have made this species an economic pillar and cornerstone of Mediterranean agriculture. However, major questions about the domestication history of the olive remain unanswered. New paleobotanical, archeological, historical and molecular data have recently accumulated for olive, making it timely to carry out a critical re-evaluation of the biogeography of wild olives and the history of their cultivation. We review here the chronological history of wild olives and discuss the questions that remain unanswered, or even unasked, about their domestication history in the Mediterranean Basin. We argue that more detailed ecological genomics studies of wild and cultivated olives are crucial to improve our understanding of olive domestication. Multidisciplinary research integrating genomics, metagenomics and community ecology will make it possible to decipher the evolutionary ecology of one of the most iconic domesticated fruit trees worldwide.
Conclusion: The olive is a relevant model for improving our knowledge of domestication processes in clonally propagated perennial crops, particularly those of the Mediterranean Basin. Future studies on the ecological and genomic shifts linked to domestication in olive and its associated community will provide insight into the phenotypic and molecular bases of crop adaptation to human uses.
© The Author(s) 2017. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
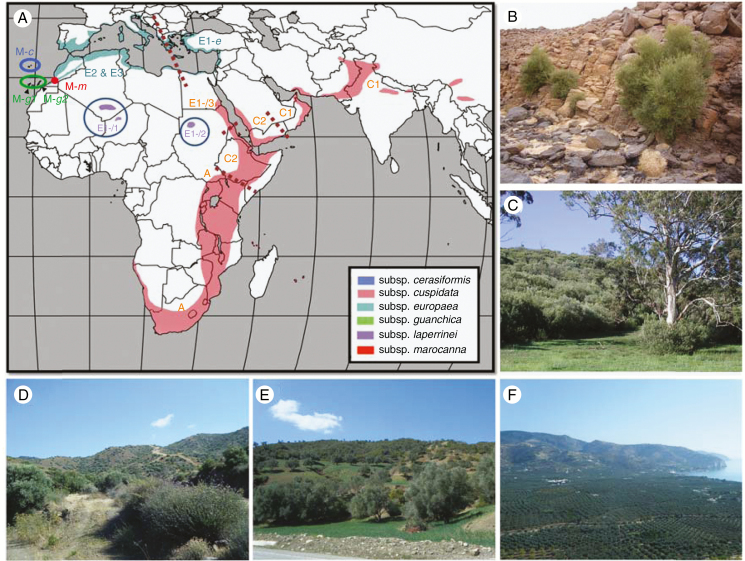
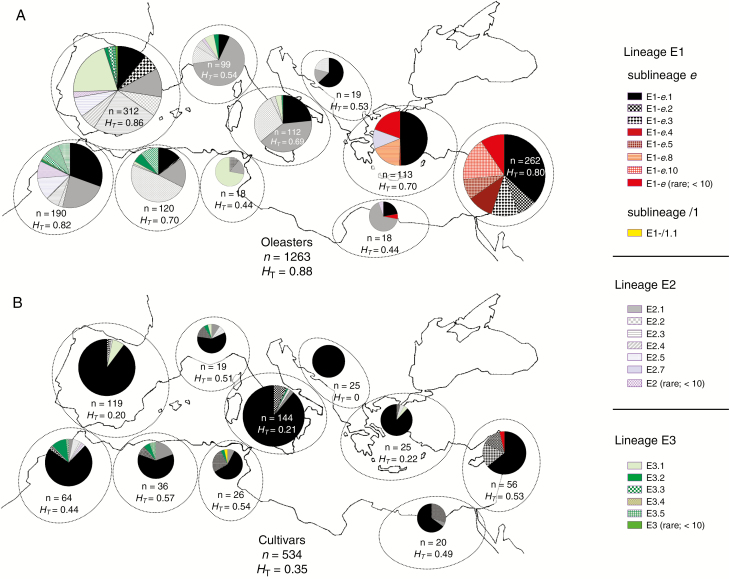
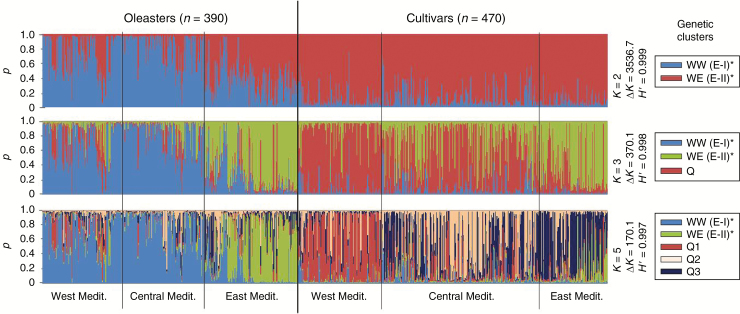
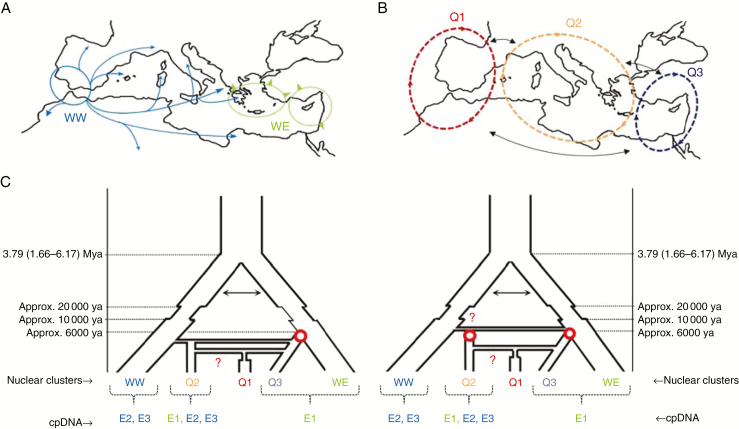
References
-
- Albertini E, Torricelli R, Bitocchi E et al. 2011. Structure of genetic diversity in Olea europaea L. cultivars from central Italy. Molecular Breeding 27: 533–547.
-
- Alcántara JM, Rey PJ. 2003. Conflicting selection pressures on seed size: evolutionary ecology of fruit size in a bird-dispersed tree, Olea europaea. Journal of Evolutionary Biology 16: 1168–1176. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
