

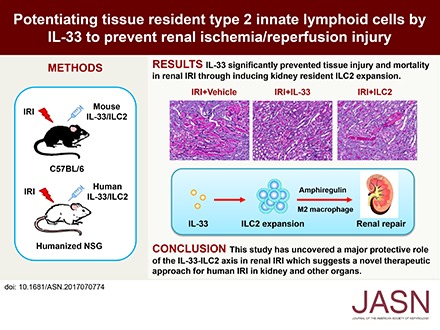
Potentiating Tissue-Resident Type 2 Innate Lymphoid Cells by IL-33 to Prevent Renal Ischemia-Reperfusion Injury
- PMID: 29295873
- PMCID: PMC5827602
- DOI: 10.1681/ASN.2017070774
Potentiating Tissue-Resident Type 2 Innate Lymphoid Cells by IL-33 to Prevent Renal Ischemia-Reperfusion Injury
Abstract
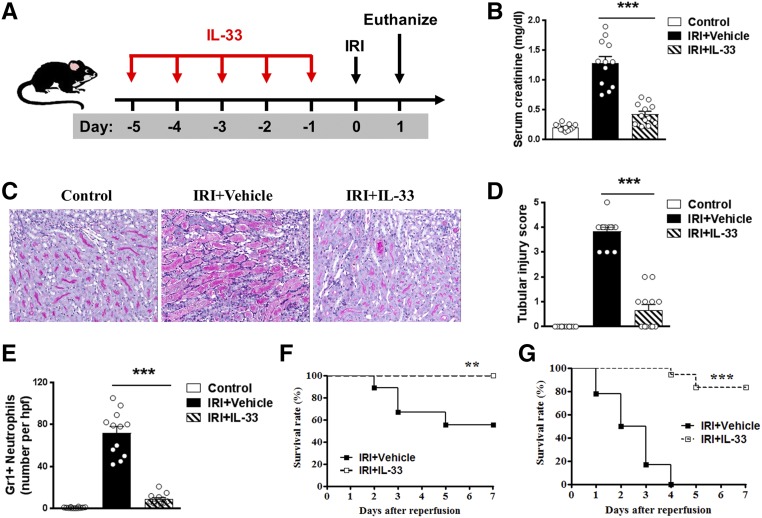
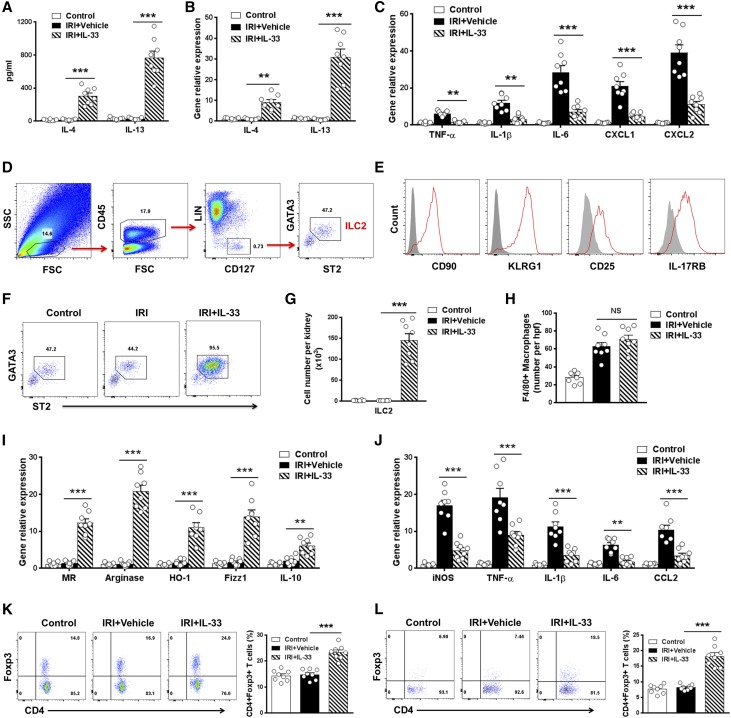
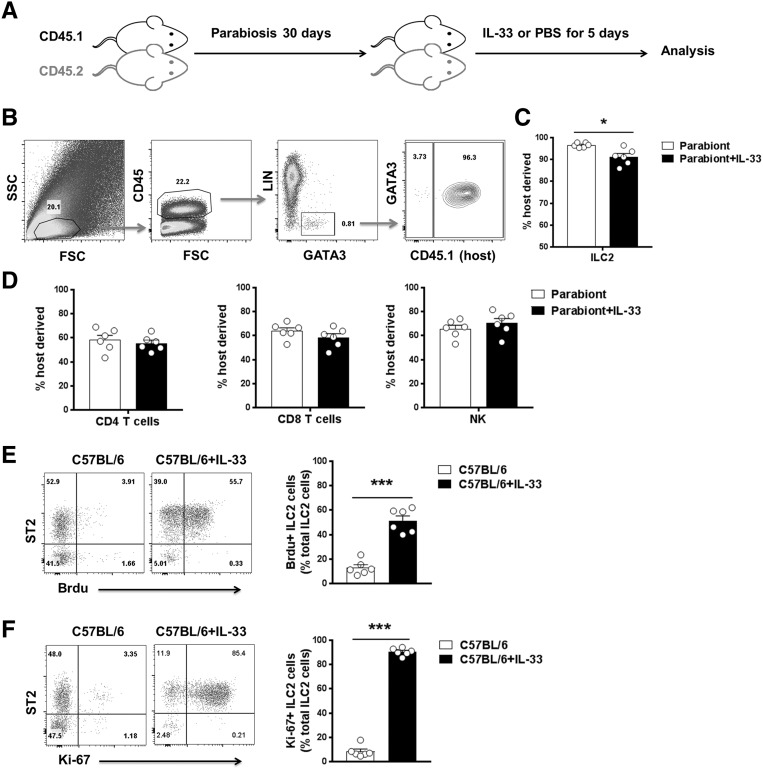
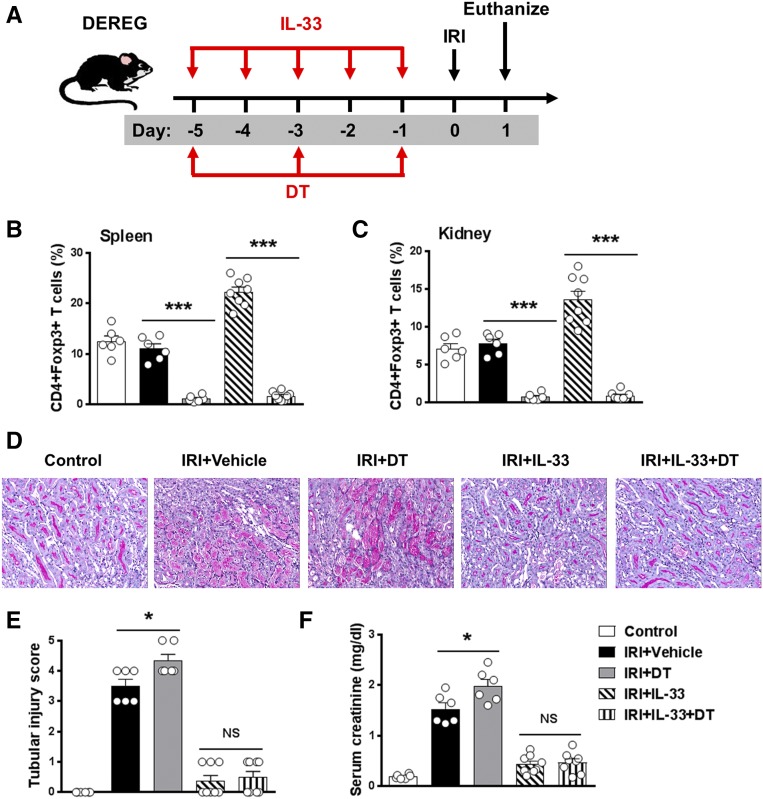
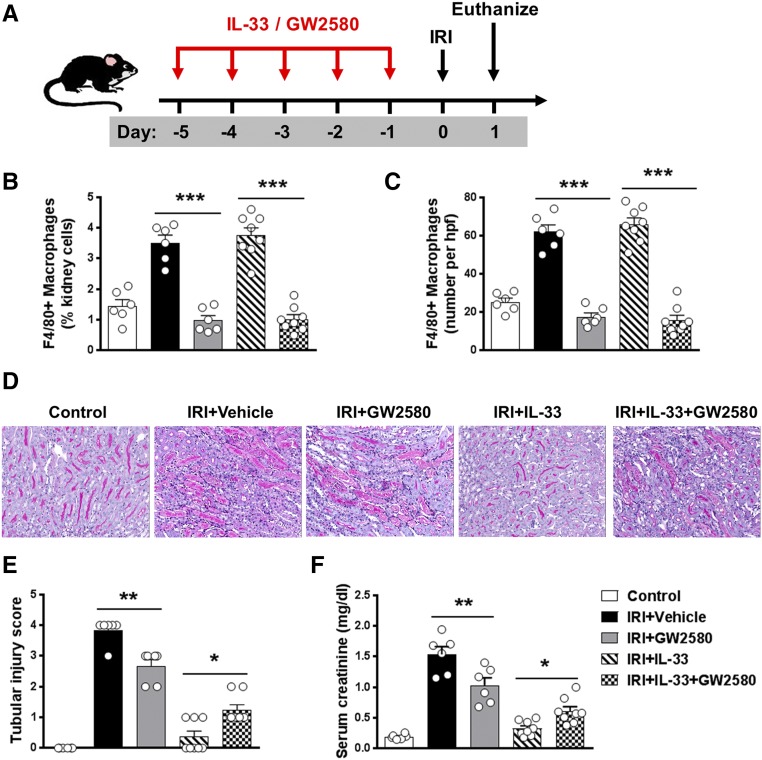
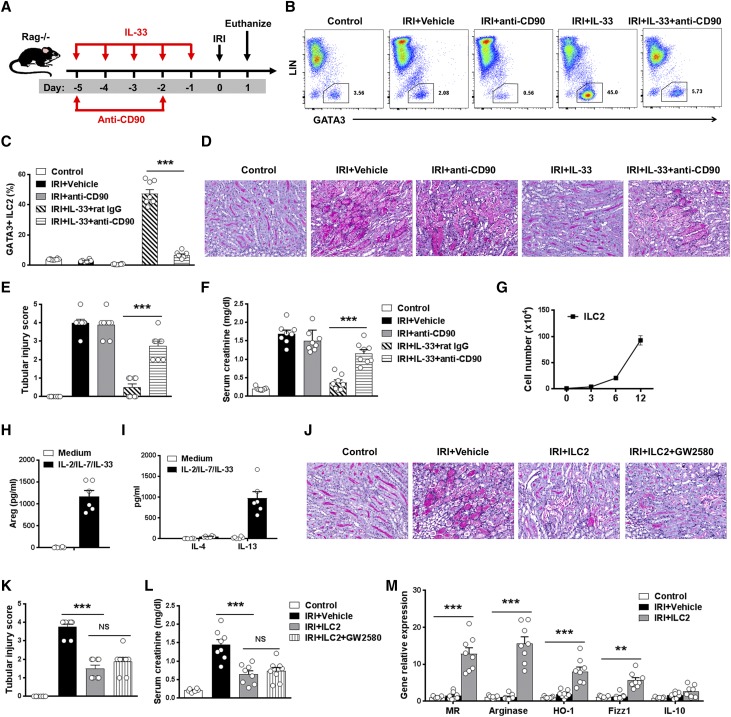
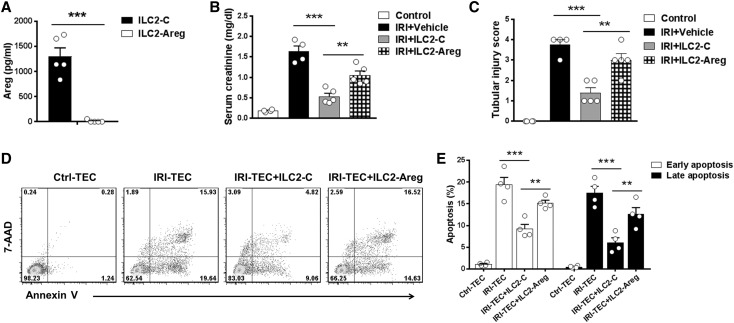
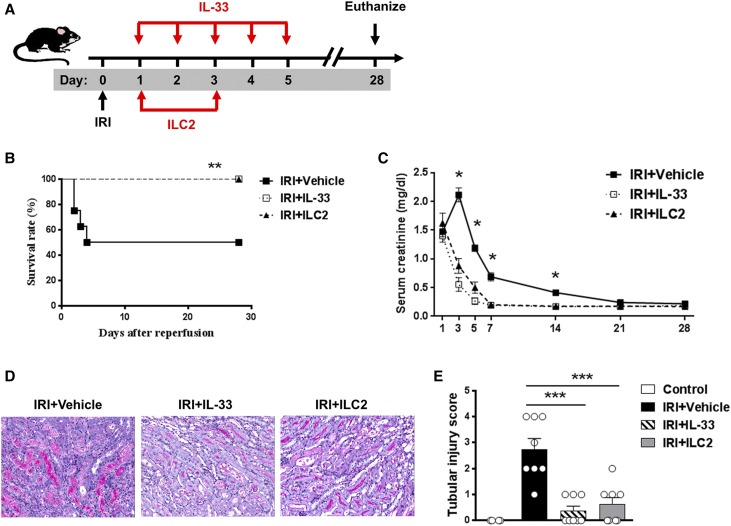
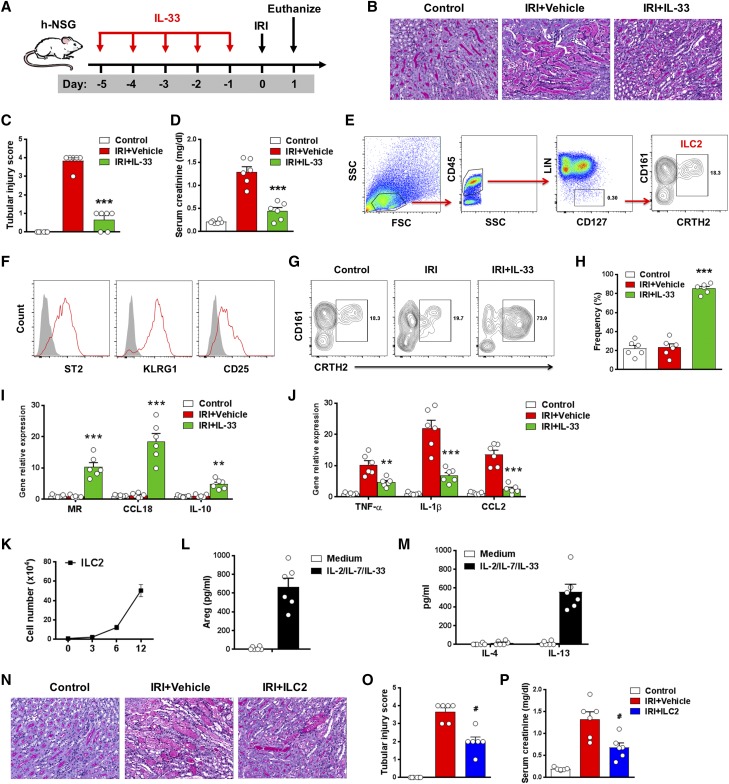
The IL-33-type 2 innate lymphoid cell (ILC2) axis has an important role in tissue homeostasis, inflammation, and wound healing. However, the relative importance of this innate immune pathway for immunotherapy against inflammation and tissue damage remains unclear. Here, we show that treatment with recombinant mouse IL-33 prevented renal structural and functional injury and reduced mortality in mice subjected to ischemia-reperfusion injury (IRI). Compared with control-treated IRI mice, IL-33-treated IRI mice had increased levels of IL-4 and IL-13 in serum and kidney and more ILC2, regulatory T cells (Tregs), and anti-inflammatory (M2) macrophages. Depletion of ILC2, but not Tregs, substantially abolished the protective effect of IL-33 on renal IRI. Adoptive transfer of ex vivo-expanded ILC2 prevented renal injury in mice subjected to IRI. This protective effect associated with induction of M2 macrophages in kidney and required ILC2 production of amphiregulin. Treatment of mice with IL-33 or ILC2 after IRI was also renoprotective. Furthermore, in a humanized mouse model of renal IRI, treatment with human IL-33 or transfer of ex vivo-expanded human ILC2 ameliorated renal IRI. This study has uncovered a major protective role of the IL-33-ILC2 axis in renal IRI that could be potentiated as a therapeutic strategy.
Keywords: IL-33; immunotherapy; innate lymphoid cells; ischemia/reperfusion injury; macrophages.
Copyright © 2018 by the American Society of Nephrology.
Figures
References
-
- Eberl G, Di Santo JP, Vivier E: The brave new world of innate lymphoid cells. Nat Immunol 16: 1–5, 2015 - PubMed
-
- Monticelli LA, Sonnenberg GF, Abt MC, Alenghat T, Ziegler CG, Doering TA, Angelosanto JM, Laidlaw BJ, Yang CY, Sathaliyawala T, Kubota M, Turner D, Diamond JM, Goldrath AW, Farber DL, Collman RG, Wherry EJ, Artis D: Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat Immunol 12: 1045–1054, 2011 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
