

Vacuolar Transporters - Companions on a Longtime Journey
- PMID: 29295940
- PMCID: PMC5813537
- DOI: 10.1104/pp.17.01481
Vacuolar Transporters - Companions on a Longtime Journey
Abstract
Biochemical and electrophysiological studies on plant vacuolar transporters became feasible in the late 1970s and early 1980s, when methods to isolate large quantities of intact vacuoles and purified vacuolar membrane vesicles were established. However, with the exception of the H+-ATPase and H+-PPase, which could be followed due to their hydrolytic activities, attempts to purify tonoplast transporters were for a long time not successful. Heterologous complementation, T-DNA insertion mutants, and later proteomic studies allowed the next steps, starting from the 1990s. Nowadays, our knowledge about vacuolar transporters has increased greatly. Nevertheless, there are several transporters of central importance that have still to be identified at the molecular level or have even not been characterized biochemically. Furthermore, our knowledge about regulation of the vacuolar transporters is very limited, and much work is needed to get a holistic view about the interplay of the vacuolar transportome. The huge amount of information generated during the last 35 years cannot be summarized in such a review. Therefore, I decided to concentrate on some aspects where we were involved during my research on vacuolar transporters, for some our laboratories contributed more, while others contributed less.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures
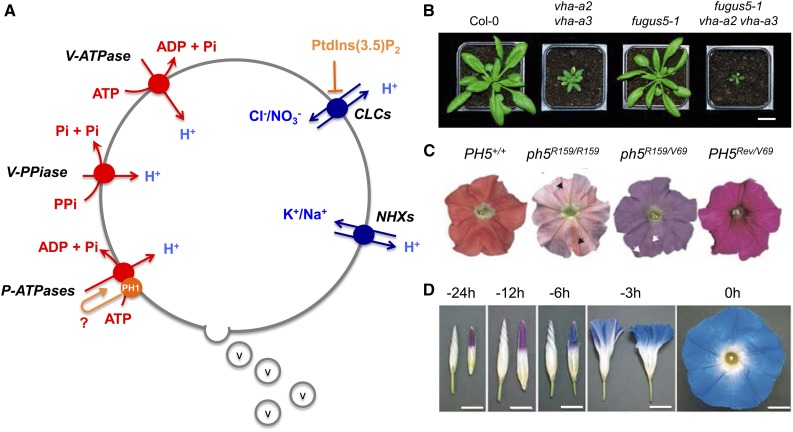
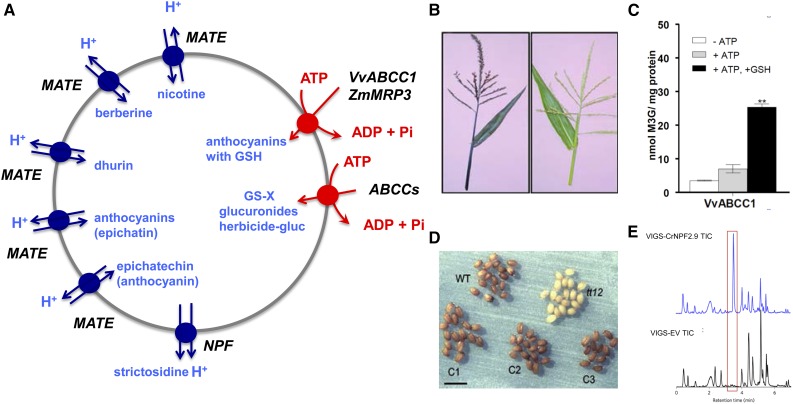
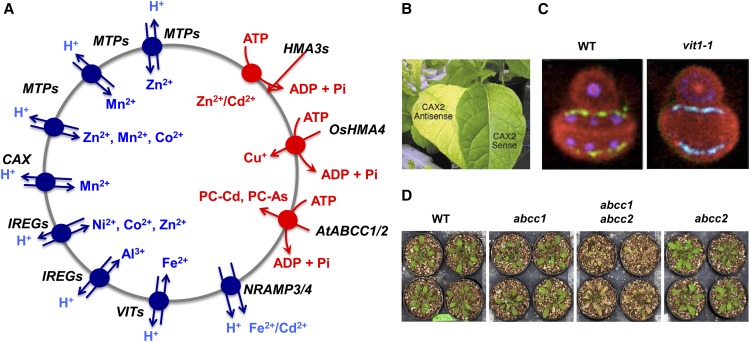
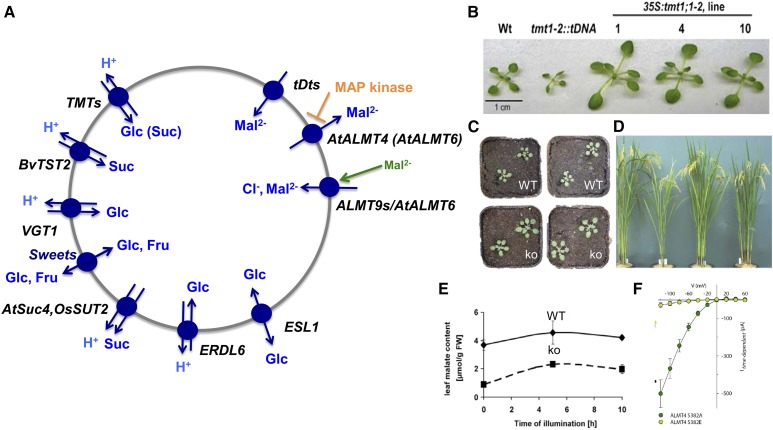
References
-
- Allen GJ, Chu SP, Schumacher K, Shimazaki CT, Vafeados D, Kemper A, Hawke SD, Tallman G, Tsien RY, Harper JF, et al. (2000) Alteration of stimulus-specific guard cell calcium oscillations and stomatal closing in Arabidopsis det3 mutant. Science 289: 2338–2342 - PubMed
-
- Appelhagen I, Nordholt N, Seidel T, Spelt K, Koes R, Quattrocchio F, Sagasser M, Weisshaar B (2015) TRANSPARENT TESTA 13 is a tonoplast P3A-ATPase required for vacuolar deposition of proanthocyanidins in Arabidopsis thaliana seeds. Plant J 82: 840–849 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
