

Inhibition of Poly(A)-binding protein with a synthetic RNA mimic reduces pain sensitization in mice
- PMID: 29295980
- PMCID: PMC5750225
- DOI: 10.1038/s41467-017-02449-5
Inhibition of Poly(A)-binding protein with a synthetic RNA mimic reduces pain sensitization in mice
Abstract
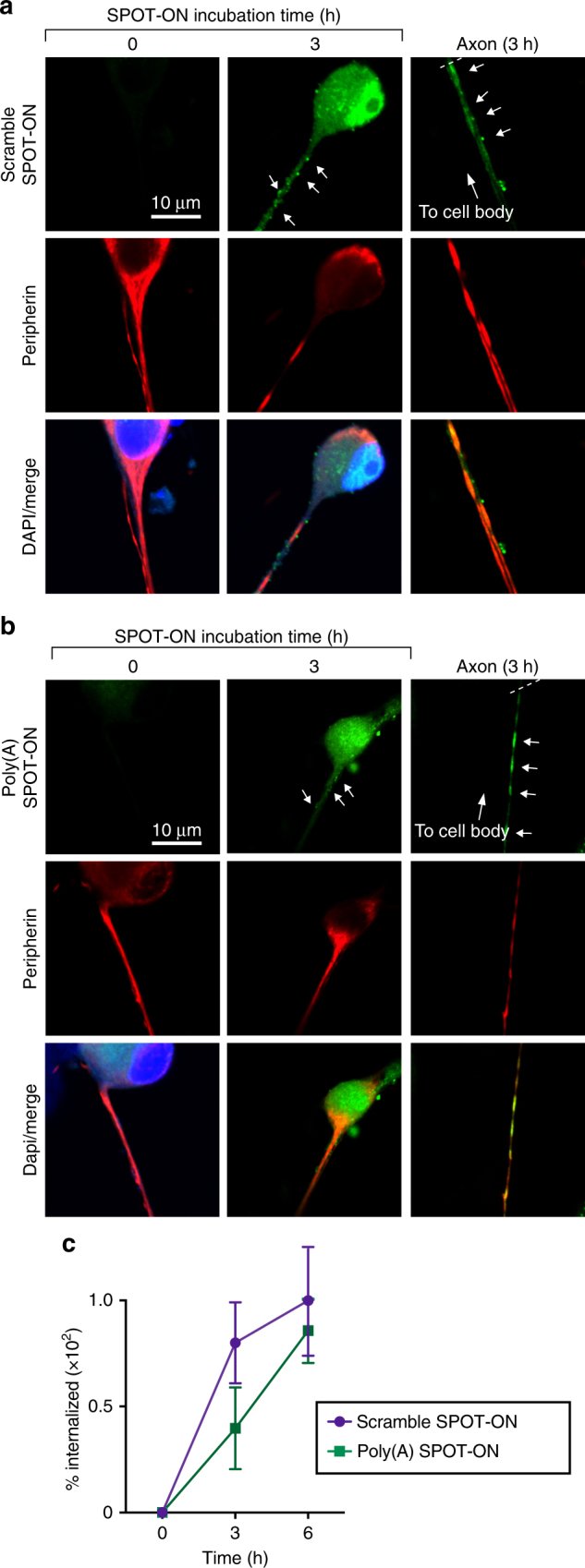
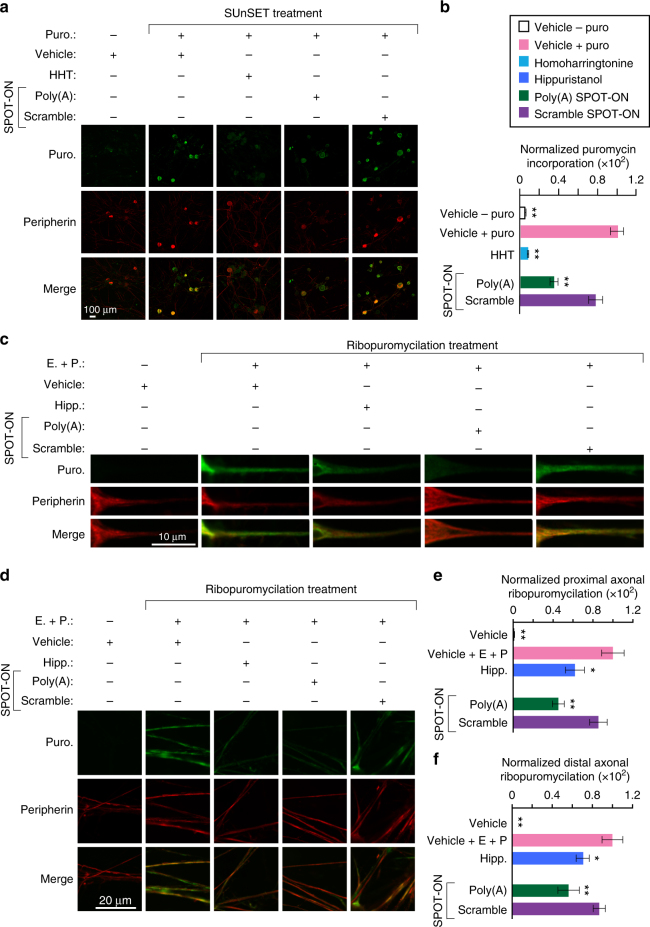
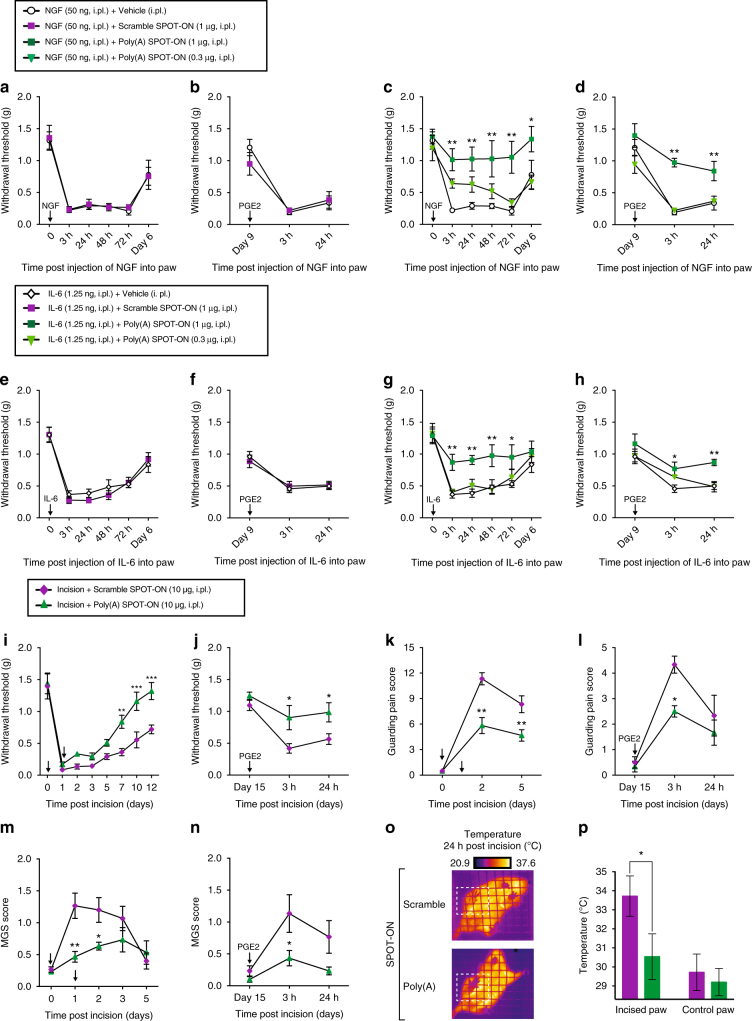
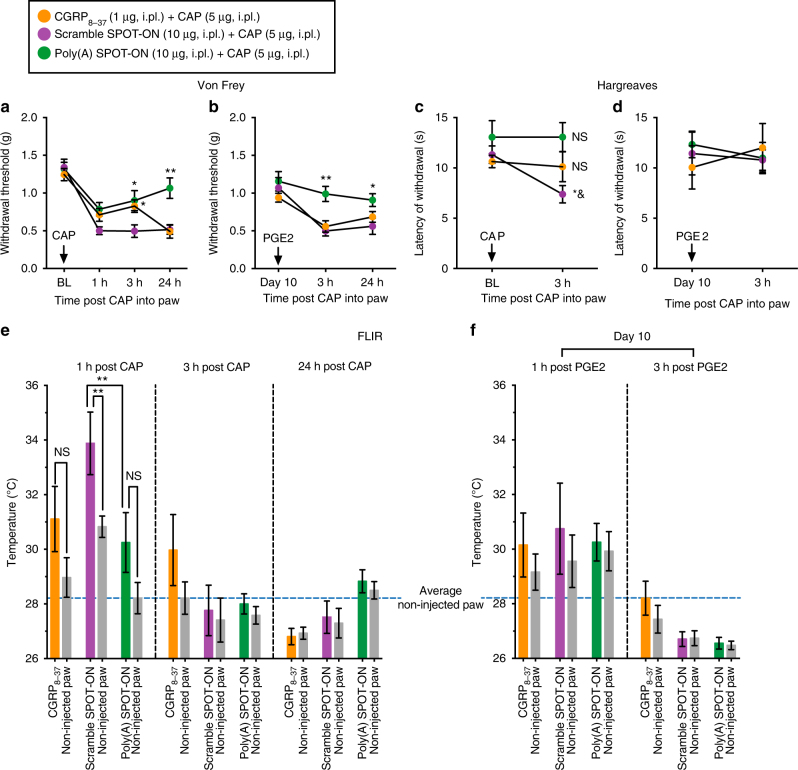
Nociceptors rely on cap-dependent translation to rapidly induce protein synthesis in response to pro-inflammatory signals. Comparatively little is known regarding the role of the regulatory factors bound to the 3' end of mRNA in nociceptor sensitization. Poly(A)-binding protein (PABP) stimulates translation initiation by bridging the Poly(A) tail to the eukaryotic initiation factor 4F complex associated with the mRNA cap. Here, we use unbiased assessment of PABP binding specificity to generate a chemically modified RNA-based competitive inhibitor of PABP. The resulting RNA mimic, which we designated as the Poly(A) SPOT-ON, is more stable than unmodified RNA and binds PABP with high affinity and selectivity in vitro. We show that injection of the Poly(A) SPOT-ON at the site of an injury can attenuate behavioral response to pain. Collectively, these results suggest that PABP is integral for nociceptive plasticity. The general strategy described here provides a broad new source of mechanism-based inhibitors for RNA-binding proteins and is applicable for in vivo studies.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
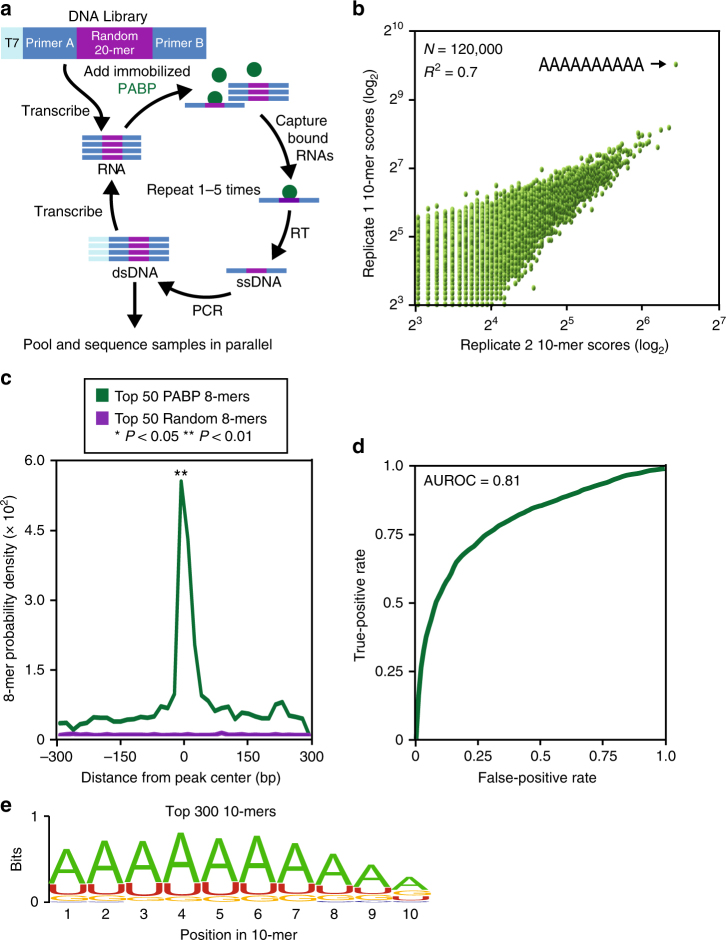
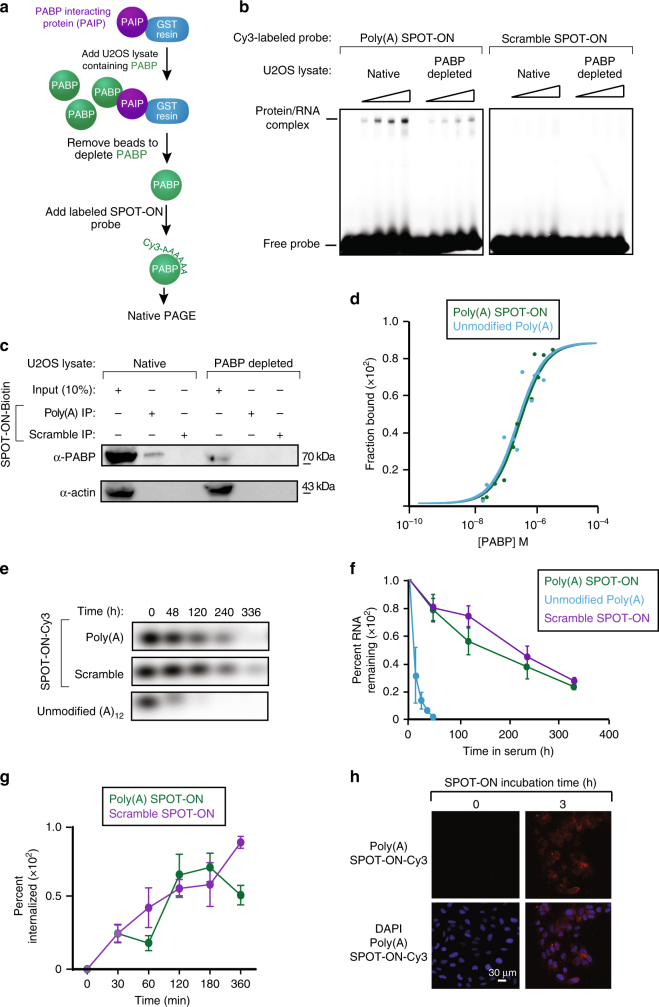
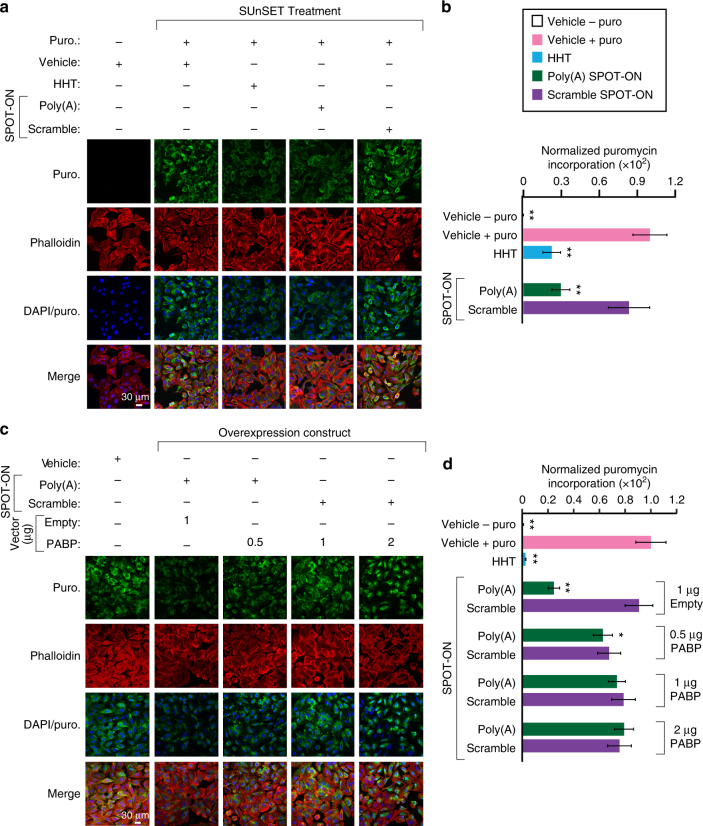
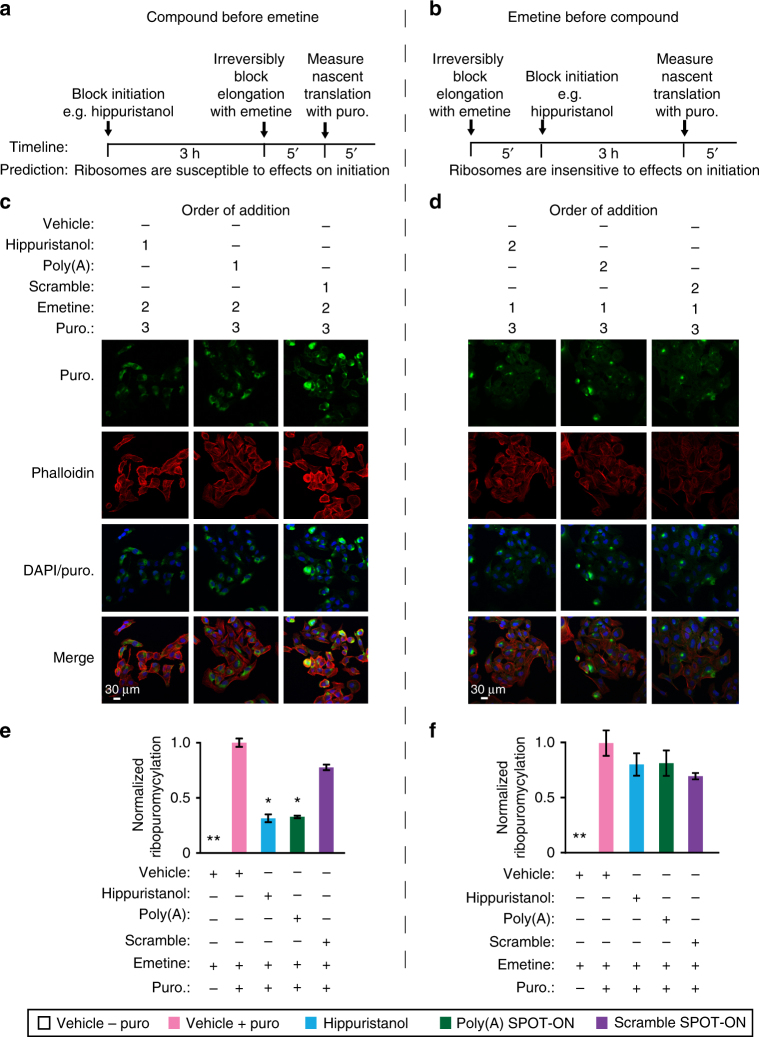

References
-
- Wang H, Tiedge H. Translational control at the synapse. Neurosci. Rev. J. Bringing Neurobiol. Neurol. Psychiatry. 2004;10:456–466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
