

IL-33, IL-25, and TSLP induce a distinct phenotypic and activation profile in human type 2 innate lymphoid cells
- PMID: 29296700
- PMCID: PMC5728348
- DOI: 10.1182/bloodadvances.2016002352
IL-33, IL-25, and TSLP induce a distinct phenotypic and activation profile in human type 2 innate lymphoid cells
Abstract
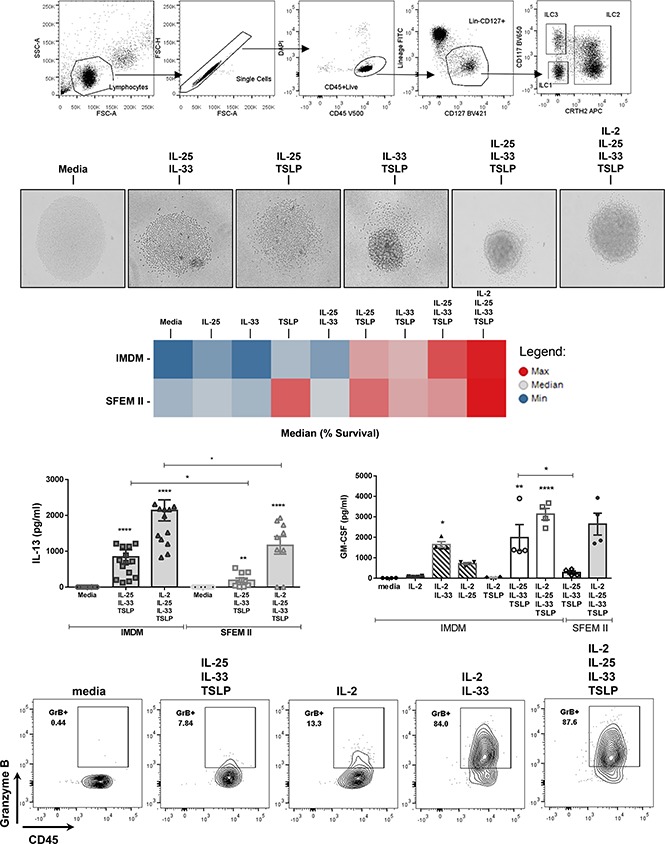
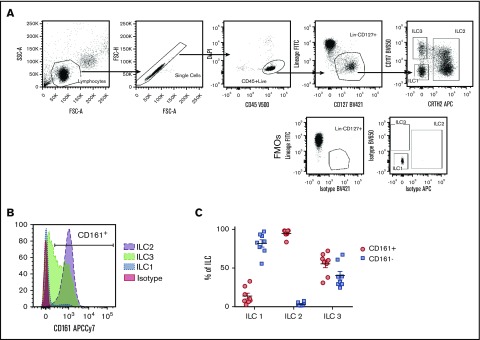
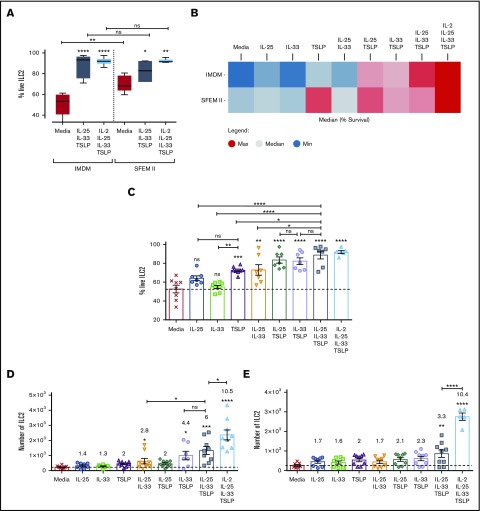
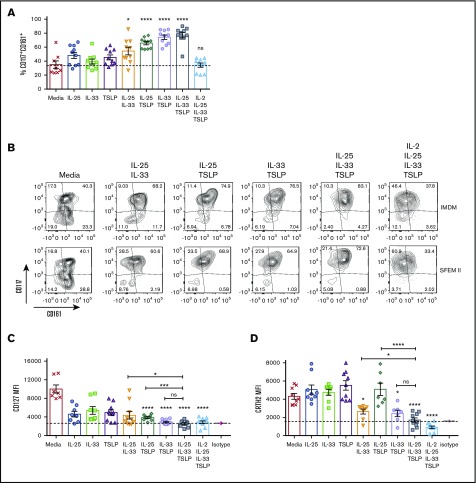
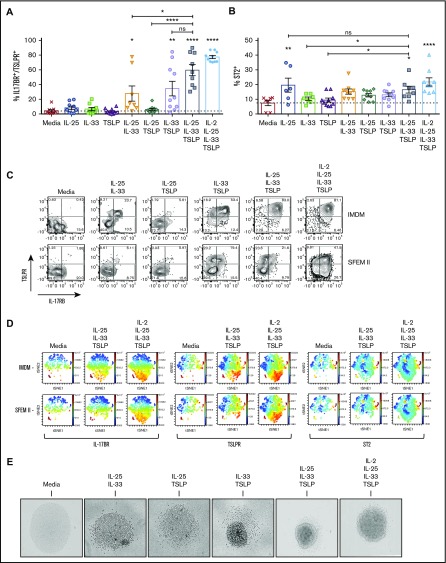
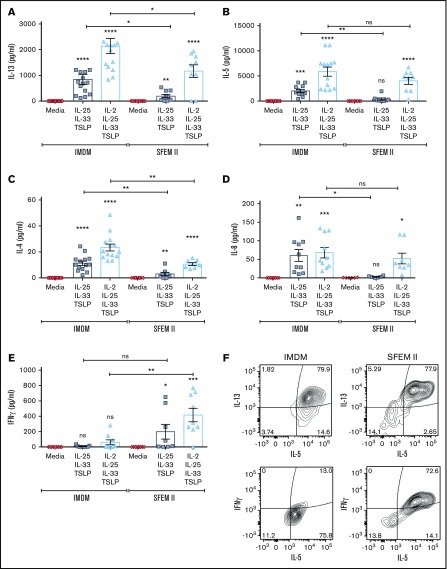
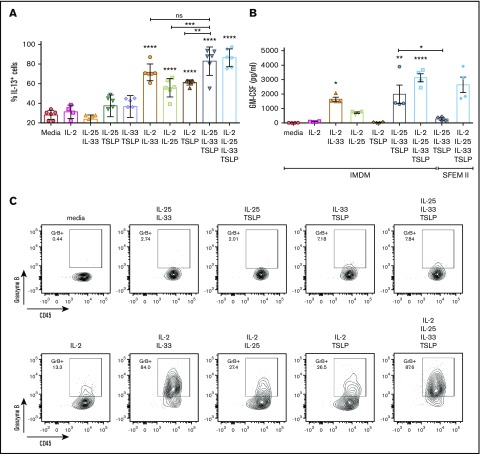
Innate lymphoid cells (ILCs) represent a distinct branch of the lymphoid lineage composed of 3 major subpopulations: ILC1, ILC2, and ILC3. ILCs are mainly described as tissue-resident cells but can be detected at low levels in human blood. However, unlike mouse ILCs, there is still no consistent methodology to purify and culture these cells that enables in-depth analysis of their intrinsic biology. Here, we describe defined culture conditions for ILC2s, which allowed us to dissect the roles of interleukin 2 (IL-2), IL-25, IL-33, and thymic stromal lymphopoietin (TSLP) individually, or in combination, in modulating ILC2 phenotype and function. We show that TSLP is important for ILC2 survival, while ILC2 activation is more dependent on IL-33, especially when in combination with IL-2 or TSLP. We found that activation of ILC2s by IL-33 and TSLP dramatically upregulated their surface expression of c-Kit and downregulated expression of the canonical markers IL-7Rα and CRTH2. IL-2 further amplified ILC2 production of IL-5, IL-13, and granulocyte-macrophage colony-stimulating factor but also induced a more natural killer (NK)-like phenotype in ILC2, with upregulation of granzyme B production by these cells. Furthermore, ILC2 plasticity was observed in serum-free SFEM II media in response to IL-33, IL-25, and TSLP stimulation and independently of IL-12 and IL-1β. This is the first comprehensive report of an in vitro culture system for human ILC2s, without the use of feeder layers, which additionally evaluates the impact of IL-25, IL-33, and TSLP alone or in combination on ILC2 surface phenotype and activation status.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
References
-
- Klose CS, Artis D. Innate lymphoid cells as regulators of immunity, inflammation and tissue homeostasis. Nat Immunol. 2016;17(7):765-774. - PubMed
-
- Barlow JL, Peel S, Fox J, et al. IL-33 is more potent than IL-25 in provoking IL-13-producing nuocytes (type 2 innate lymphoid cells) and airway contraction. J Allergy Clin Immunol. 2013;132(4):933-941. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
