

A functional DC cross talk promotes human ILC homeostasis in humanized mice
- PMID: 29296702
- PMCID: PMC5728352
- DOI: 10.1182/bloodadvances.2017004358
A functional DC cross talk promotes human ILC homeostasis in humanized mice
Abstract
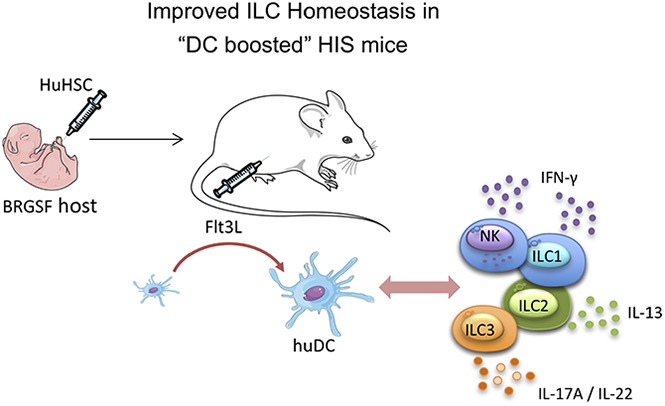
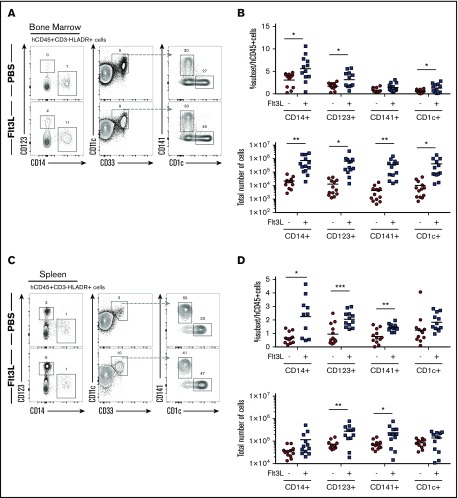
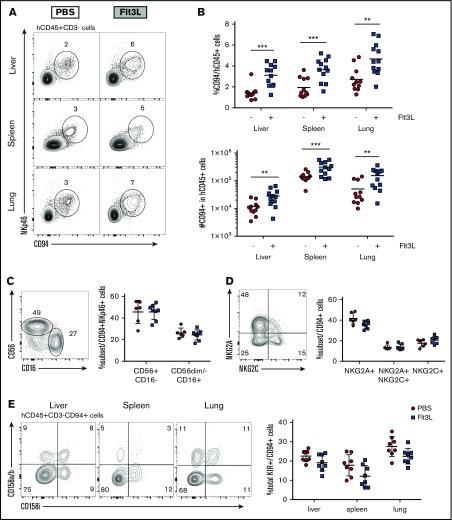
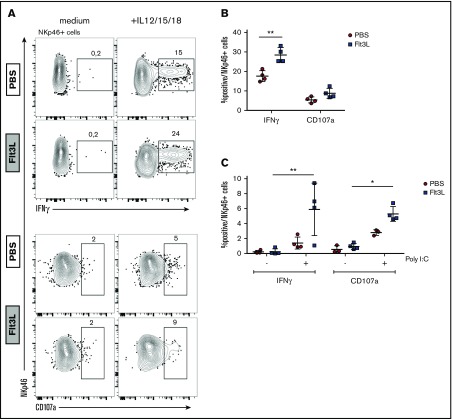
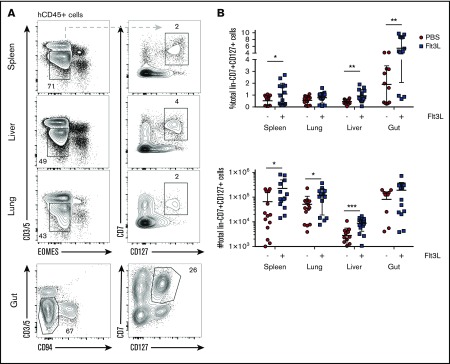
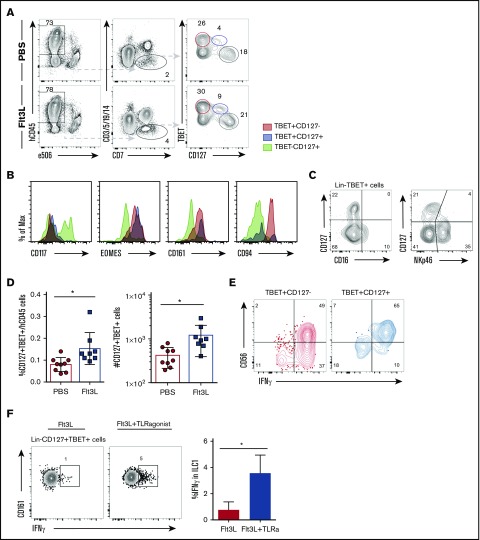
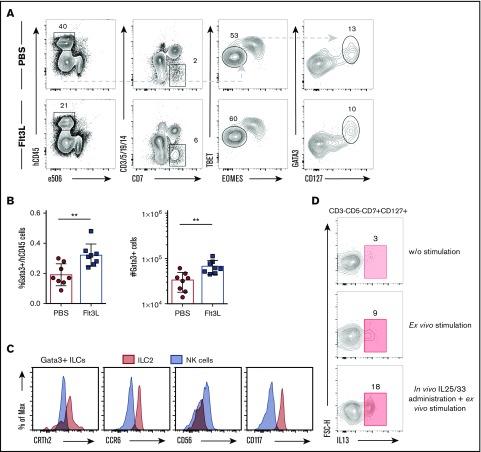
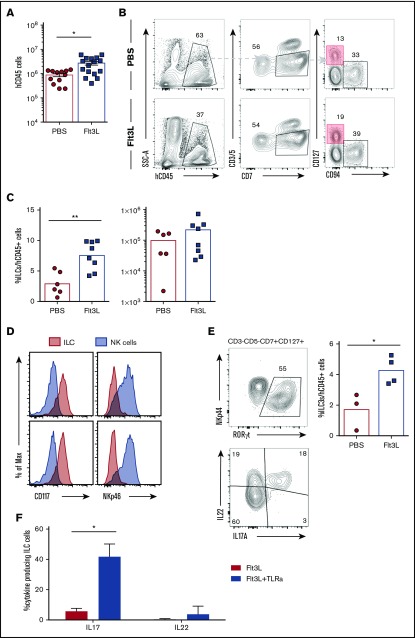
Humanized mice harboring human hematopoietic systems offer a valuable small-animal model to assess human immune responses to infection, inflammation, and cancer. Human immune system (HIS) mice develop a broad repertoire of antigen receptor bearing B and T cells that can participate in adaptive immune responses after immunization. In contrast, analysis of innate immune components, including innate lymphoid cells (ILCs) and natural killer (NK) cells, is limited in current HIS mouse models, partly because of the poor development of these rare lymphoid subsets. Here we show that novel dendritic cell (DC)-boosted BALB/c Rag2-/-Il2rg-/-SirpaNODFlk2-/- (BRGSF) HIS mice harbor abundant NK cells and tissue-resident ILC subsets in lymphoid and nonlymphoid mucosal sites. We find that human NK cells and ILCs are phenotypically and functionally mature and provide evidence that human DC activation in BRGSF-based HIS mice can "cross talk" to human NK cells and ILCs. This novel HIS mouse model should provide the opportunity to study the immunobiology of human NK cell and ILC subsets in vivo in response to various environmental challenges.
Conflict of interest statement
Conflict-of-interest disclosure: J.P.D. is a stakeholder in AXENIS (founder, member of the executive board). The remaining authors declare no conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
