

The Protective Action Encoding of Serotonin Transients in the Human Brain
- PMID: 29297512
- PMCID: PMC5916372
- DOI: 10.1038/npp.2017.304
The Protective Action Encoding of Serotonin Transients in the Human Brain
Abstract
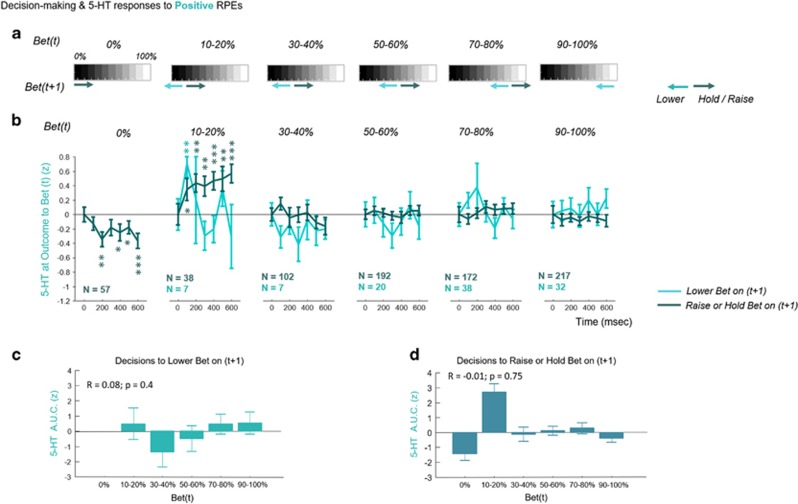
The role of serotonin in human brain function remains elusive due, at least in part, to our inability to measure rapidly the local concentration of this neurotransmitter. We used fast-scan cyclic voltammetry to infer serotonergic signaling from the striatum of 14 brains of human patients with Parkinson's disease. Here we report these novel measurements and show that they correlate with outcomes and decisions in a sequential investment game. We find that serotonergic concentrations transiently increase as a whole following negative reward prediction errors, while reversing when counterfactual losses predominate. This provides initial evidence that the serotonergic system acts as an opponent to dopamine signaling, as anticipated by theoretical models. Serotonin transients on one trial were also associated with actions on the next trial in a manner that correlated with decreased exposure to poor outcomes. Thus, the fluctuations observed for serotonin appear to correlate with the inhibition of over-reactions and promote persistence of ongoing strategies in the face of short-term environmental changes. Together these findings elucidate a role for serotonin in the striatum, suggesting it encodes a protective action strategy that mitigates risk and modulates choice selection particularly following negative environmental events.
Figures




References
-
- Balasubramani PP, Chakravarthy VS, Ravindran B, Moustafa AA (2014). An extended reinforcement learning model of basal ganglia to understand the contributions of serotonin and dopamine in risk-based decision making, reward prediction, and punishment learning. Front Comput Neurosci 8: 47. - PMC - PubMed
-
- Chiu PH, Lohrenz TM, Montague PR (2008). Smokers' brains compute, but ignore, a fictive error signal in a sequential investment task. Nat Neurosci 11: 514–520. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
