

Interrogating the Spatiotemporal Landscape of Neuromodulatory GPCR Signaling by Real-Time Imaging of cAMP in Intact Neurons and Circuits
- PMID: 29298426
- PMCID: PMC5761078
- DOI: 10.1016/j.celrep.2017.12.022
Interrogating the Spatiotemporal Landscape of Neuromodulatory GPCR Signaling by Real-Time Imaging of cAMP in Intact Neurons and Circuits
Erratum in
-
Interrogating the Spatiotemporal Landscape of Neuromodulatory GPCR Signaling by Real-Time Imaging of cAMP in Intact Neurons and Circuits.Cell Rep. 2018 Jul 24;24(4):1081-1084. doi: 10.1016/j.celrep.2018.07.031. Cell Rep. 2018. PMID: 30044975 Free PMC article. No abstract available.
Abstract
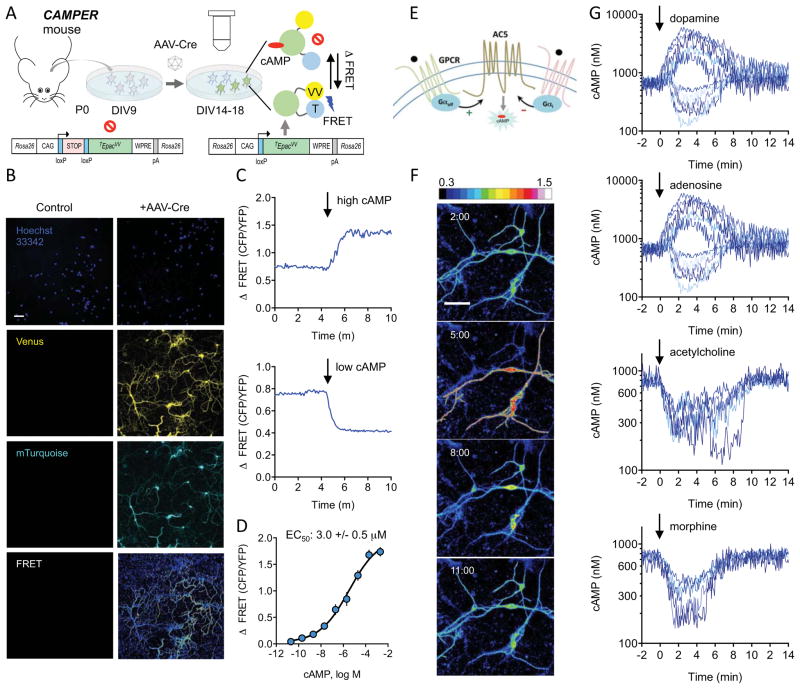
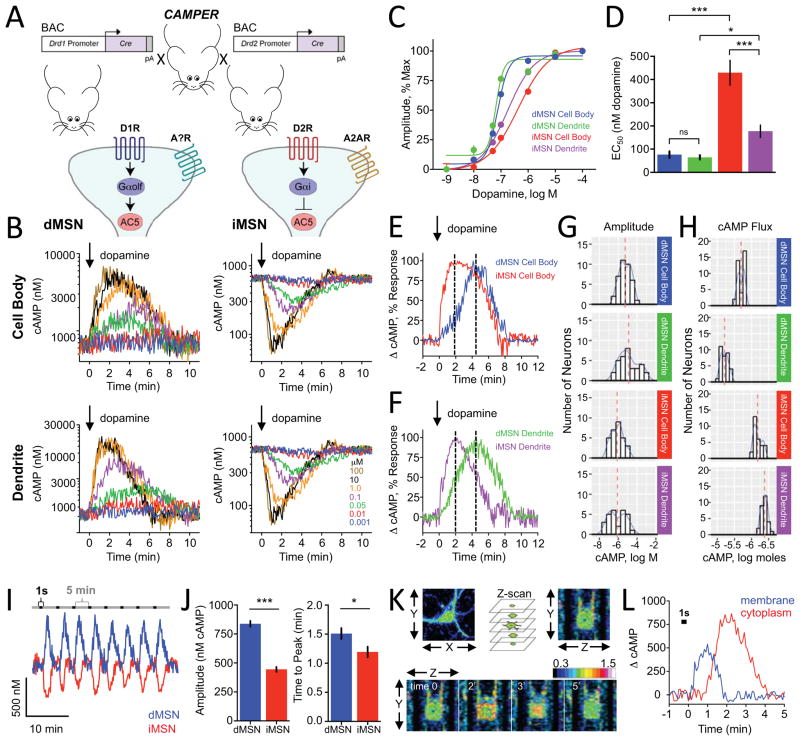
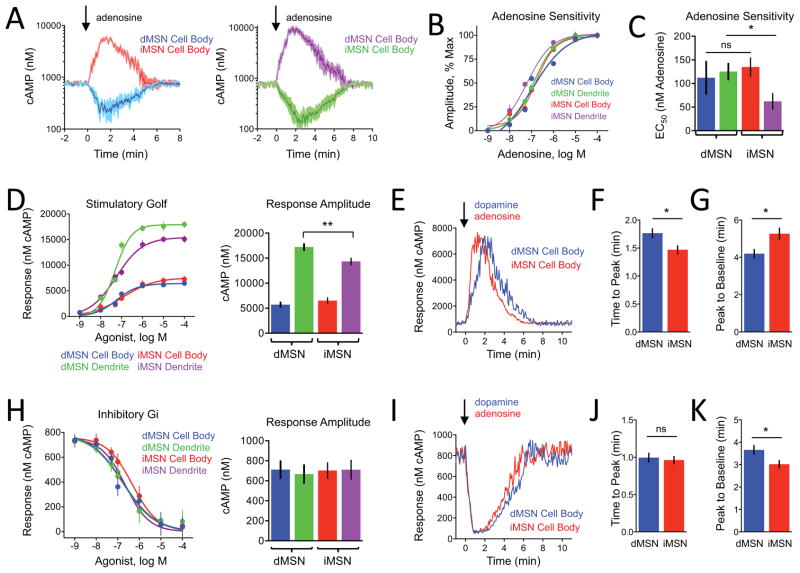
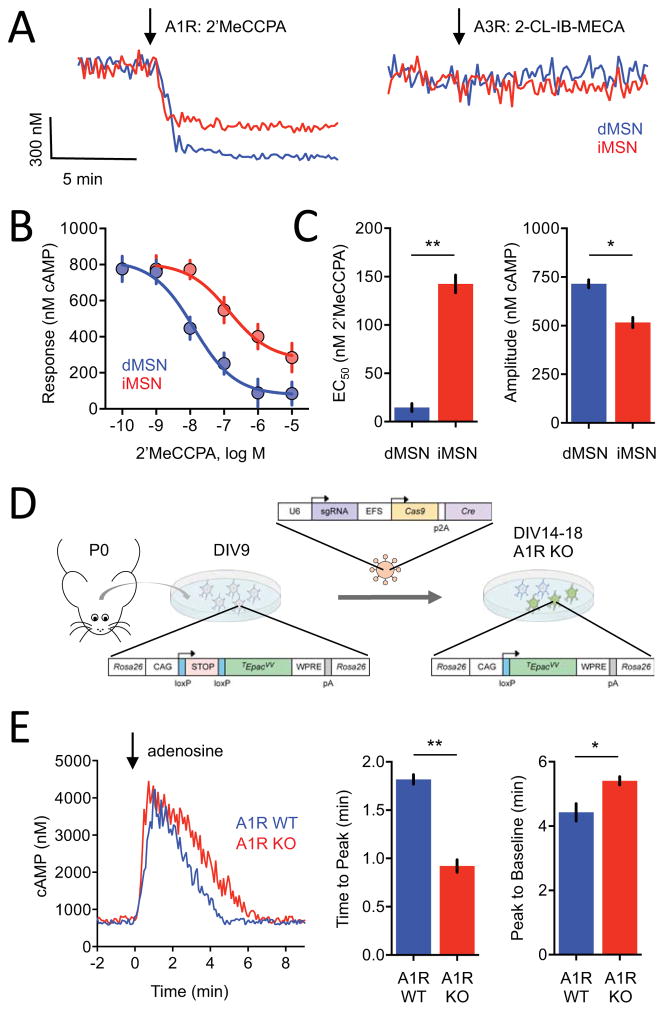
Modulation of neuronal circuits is key to information processing in the brain. The majority of neuromodulators exert their effects by activating G-protein-coupled receptors (GPCRs) that control the production of second messengers directly impacting cellular physiology. How numerous GPCRs integrate neuromodulatory inputs while accommodating diversity of incoming signals is poorly understood. In this study, we develop an in vivo tool and analytical suite for analyzing GPCR responses by monitoring the dynamics of a key second messenger, cyclic AMP (cAMP), with excellent quantitative and spatiotemporal resolution in various neurons. Using this imaging approach in combination with CRISPR/Cas9 editing and optogenetics, we interrogate neuromodulatory mechanisms of defined populations of neurons in an intact mesolimbic reward circuit and describe how individual inputs generate discrete second-messenger signatures in a cell- and receptor-specific fashion. This offers a resource for studying native neuronal GPCR signaling in real time.
Keywords: GPCR; cAMP; dopamine; imaging; neuromodulation; optogenetics; striatum.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Comment in
-
Opioid-Induced Adaptations of cAMP Dynamics in the Nucleus Accumbens.Trends Pharmacol Sci. 2020 Apr;41(4):230-232. doi: 10.1016/j.tips.2020.01.004. Epub 2020 Jan 18. Trends Pharmacol Sci. 2020. PMID: 31964511
References
-
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989;12:366–375. - PubMed
-
- Bargmann CI, Marder E. From the connectome to brain function. Nat Methods. 2013;10:483–490. - PubMed
-
- Brinton RE. Neuromodulation: associative and nonlinear adaptation. Brain research bulletin. 1990;24:651–658. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
