

Dynamic Remodeling of Pericytes In Vivo Maintains Capillary Coverage in the Adult Mouse Brain
- PMID: 29298435
- PMCID: PMC5782812
- DOI: 10.1016/j.celrep.2017.12.016
Dynamic Remodeling of Pericytes In Vivo Maintains Capillary Coverage in the Adult Mouse Brain
Abstract
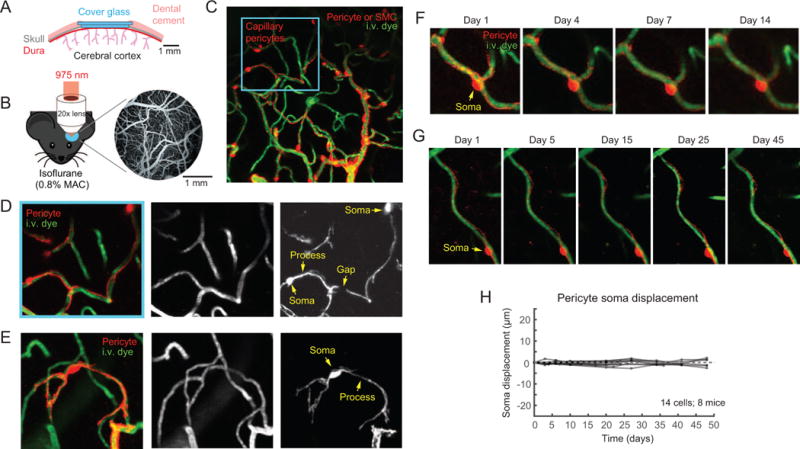
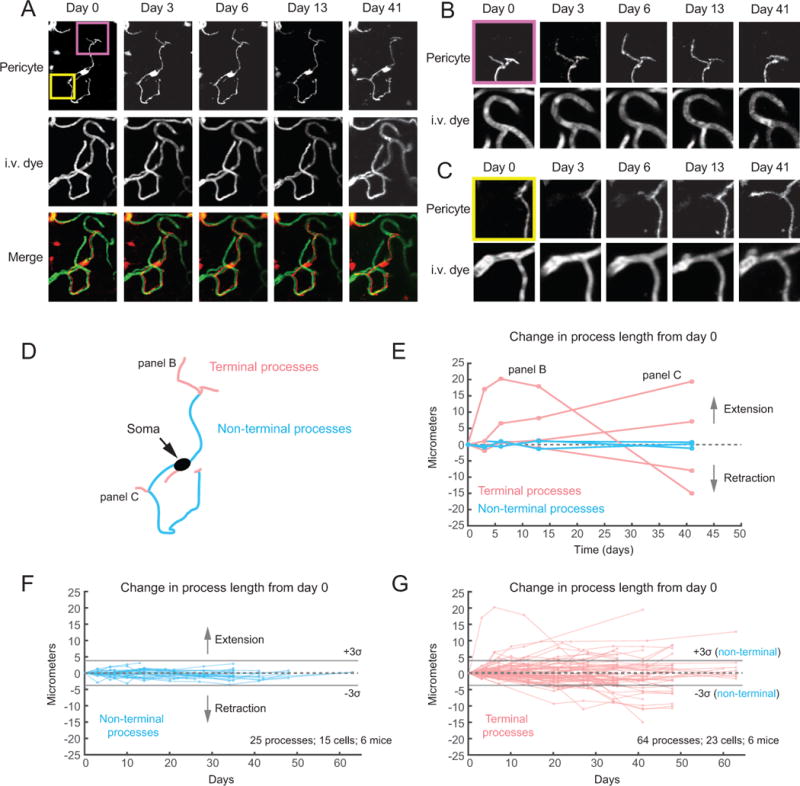
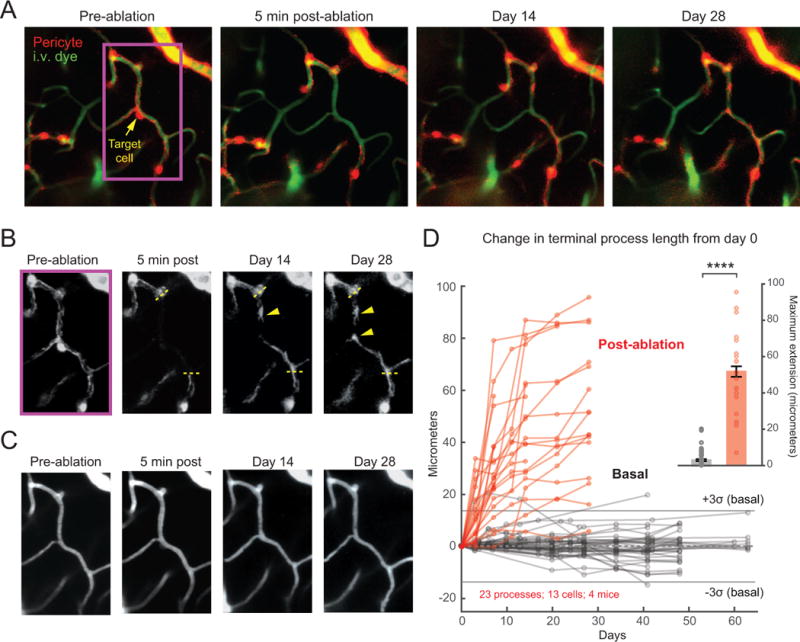
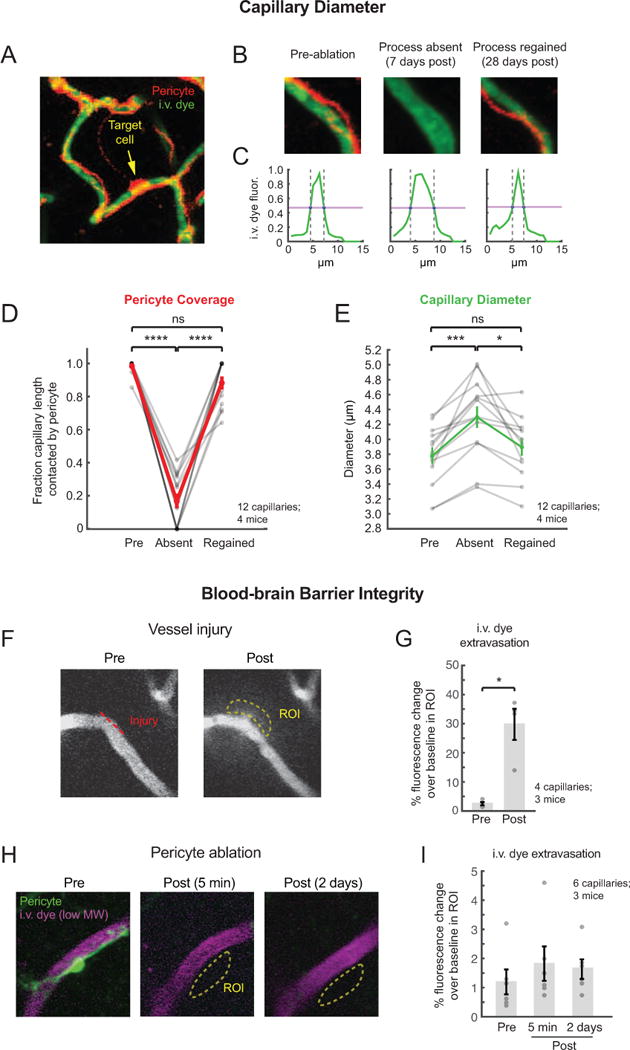
Direct contact and communication between pericytes and endothelial cells is critical for maintenance of cerebrovascular stability and blood-brain barrier function. Capillary pericytes have thin processes that reach hundreds of micrometers along the capillary bed. The processes of adjacent pericytes come in close proximity but do not overlap, yielding a cellular chain with discrete territories occupied by individual pericytes. Little is known about whether this pericyte chain is structurally dynamic in the adult brain. Using in vivo two-photon imaging in adult mouse cortex, we show that while pericyte somata were immobile, the tips of their processes underwent extensions and/or retractions over days. The selective ablation of single pericytes provoked exuberant extension of processes from neighboring pericytes to contact uncovered regions of the endothelium. Uncovered capillary regions had normal barrier function but were dilated until pericyte contact was regained. Pericyte structural plasticity may be critical for cerebrovascular health and warrants detailed investigation.
Keywords: Alzheimer’s disease; blood flow; blood-brain barrier; capillary; dementia; endothelium; pericyte; plasticity; stroke; two-photon imaging.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
References
-
- Allt G, Lawrenson JG. Pericytes: cell biology and pathology. Cells, tissue, organs. 2001;169:1–11. - PubMed
-
- Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circulation Research. 2005;97:512–523. - PubMed
-
- Armulik A, Genove G, Betsholtz C. Pericytes: Developmental, physiological, and pathological perspectives, problems and promises. Developmental Cell. 2011;21:193–215. - PubMed
-
- Armulik A, Genové G, Mäe M, Nisancioglu MH, Wallgard E, Niaudet C, He L, Norlin J, Lindblom P, Strittmatter K, et al. Pericytes regulate the blood-brain barrier. Nature. 2010;468:557–561. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
