

Mechanisms of urodele limb regeneration
- PMID: 29299322
- PMCID: PMC5743758
- DOI: 10.1002/reg2.92
Mechanisms of urodele limb regeneration
Abstract
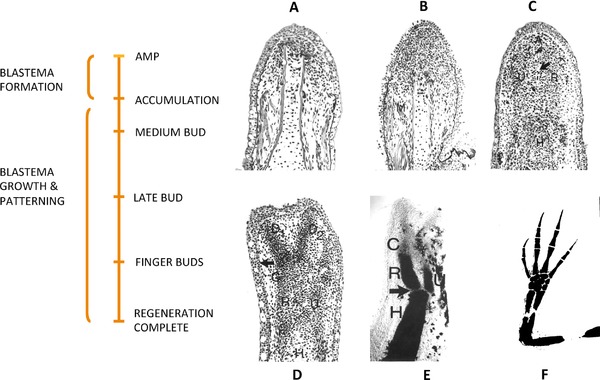
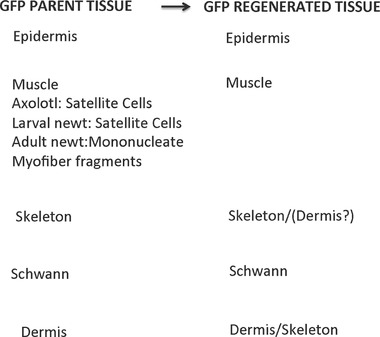
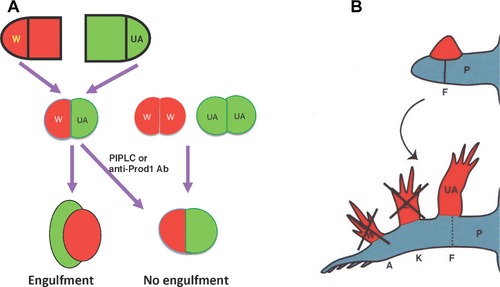
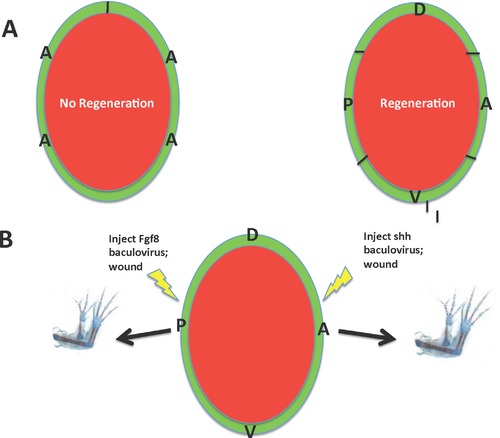
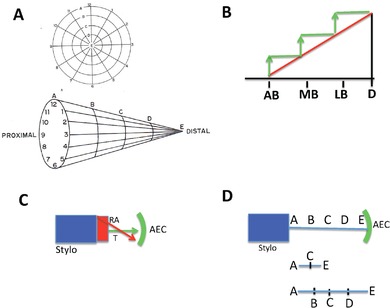
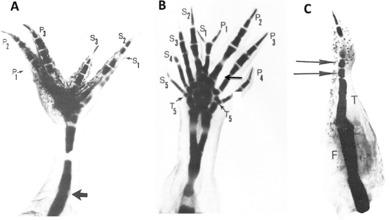
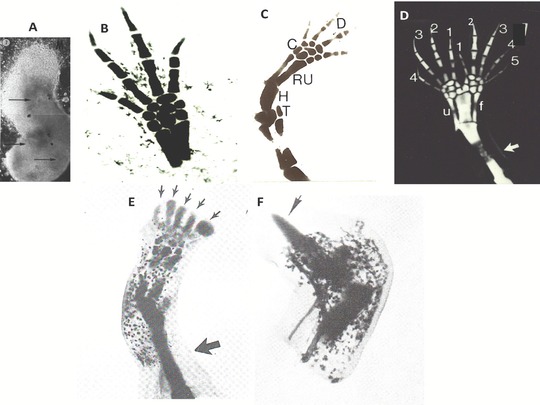
This review explores the historical and current state of our knowledge about urodele limb regeneration. Topics discussed are (1) blastema formation by the proteolytic histolysis of limb tissues to release resident stem cells and mononucleate cells that undergo dedifferentiation, cell cycle entry and accumulation under the apical epidermal cap. (2) The origin, phenotypic memory, and positional memory of blastema cells. (3) The role played by macrophages in the early events of regeneration. (4) The role of neural and AEC factors and interaction between blastema cells in mitosis and distalization. (5) Models of pattern formation based on the results of axial reversal experiments, experiments on the regeneration of half and double half limbs, and experiments using retinoic acid to alter positional identity of blastema cells. (6) Possible mechanisms of distalization during normal and intercalary regeneration. (7) Is pattern formation is a self-organizing property of the blastema or dictated by chemical signals from adjacent tissues? (8) What is the future for regenerating a human limb?
Keywords: limb; mechanisms; regeneration; review; urodele.
Figures
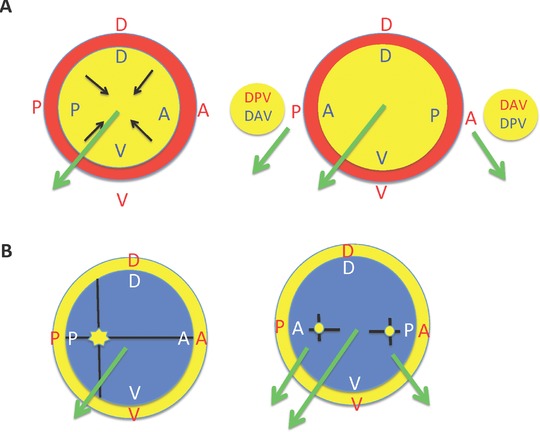
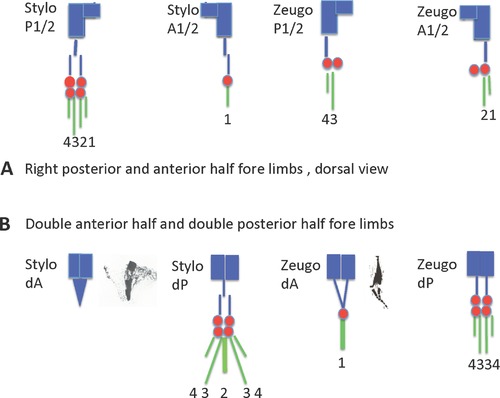
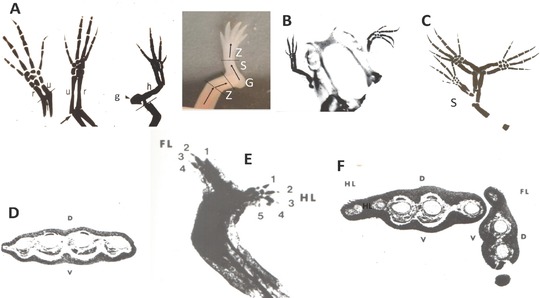
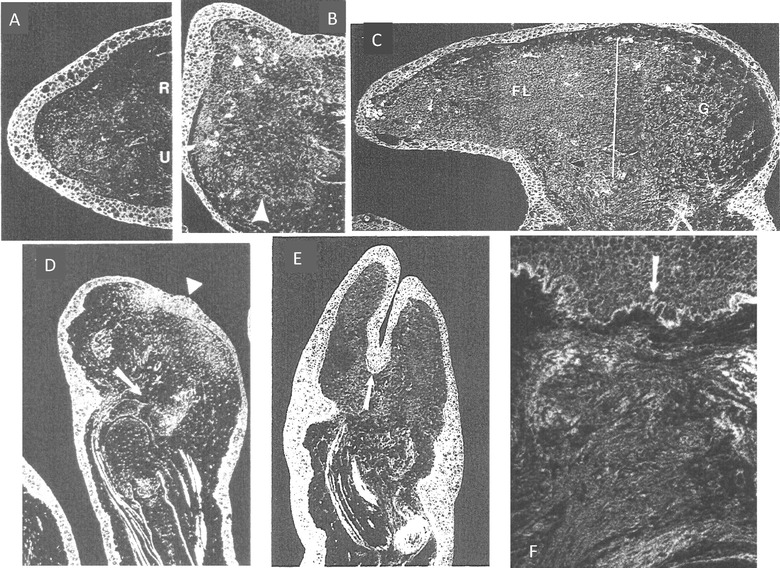
References
-
- Adams, D. S. , Masi, A. , & Levin, M. (2007). H+ pump‐dependent changes in membrane voltage are an early mechanism necessary and sufficient to induce Xenopus tail regeneration. Development, 134, 1323–1335. - PubMed
-
- Albert, P. , & Boilly, B. (1988). Effect of transferrin on amphibian limb regeneration: a blastema cell culture study. Roux's Archives for Developmental Biology, 197, 193–196. - PubMed
-
- Albert, P. , Boilly, B. , Courty, J. , & Barritault, D. (1987). Stimulation in cell culture of mesenchymal cells of newt limb blastemas by EDGFI or II (basic or acidic FGF). Cell Differentiation, 21, 63–68. - PubMed
-
- Anastasia, L. , Sampaolesi, M. , Papini, N. , Oleari, D. , Lamorte, G. , Tringali, C. , … Venerando, B . (2006). Reversine‐treated fibroblasts acquire myogenic competence in vitro and in regenerating skeletal muscle. Cell Death and Differentiation, 13, 2042–2051. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous