

Autophagy Is Required for Sortilin-Mediated Degradation of Apolipoprotein B100
- PMID: 29301854
- PMCID: PMC5815905
- DOI: 10.1161/CIRCRESAHA.117.311240
Autophagy Is Required for Sortilin-Mediated Degradation of Apolipoprotein B100
Abstract
Rationale: Genome-wide association studies identified single-nucleotide polymorphisms near the SORT1 locus strongly associated with decreased plasma LDL-C (low-density lipoprotein cholesterol) levels and protection from atherosclerotic cardiovascular disease and myocardial infarction. The minor allele of the causal SORT1 single-nucleotide polymorphism locus creates a putative C/EBPα (CCAAT/enhancer-binding protein α)-binding site in the SORT1 promoter, thereby increasing in homozygotes sortilin expression by 12-fold in liver, which is rich in this transcription factor. Our previous studies in mice have showed reductions in plasma LDL-C and its principal protein component, apoB (apolipoprotein B) with increased SORT1 expression, and in vitro studies suggested that sortilin promoted the presecretory lysosomal degradation of apoB associated with the LDL precursor, VLDL (very-low-density lipoprotein).
Objective: To determine directly that SORT1 overexpression results in apoB degradation and to identify the mechanisms by which this reduces apoB and VLDL secretion by the liver, thereby contributing to understanding the clinical phenotype of lower LDL-C levels.
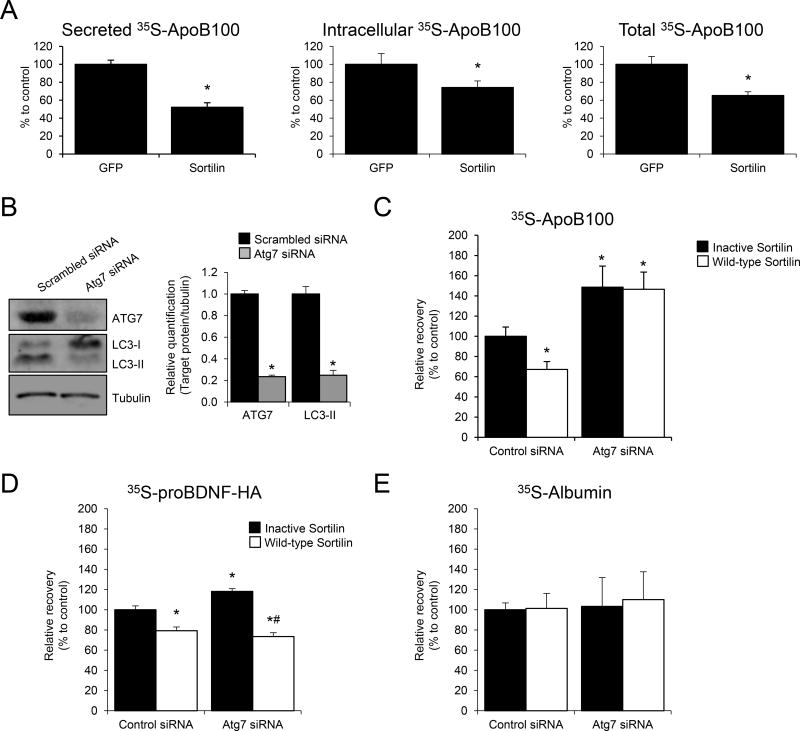
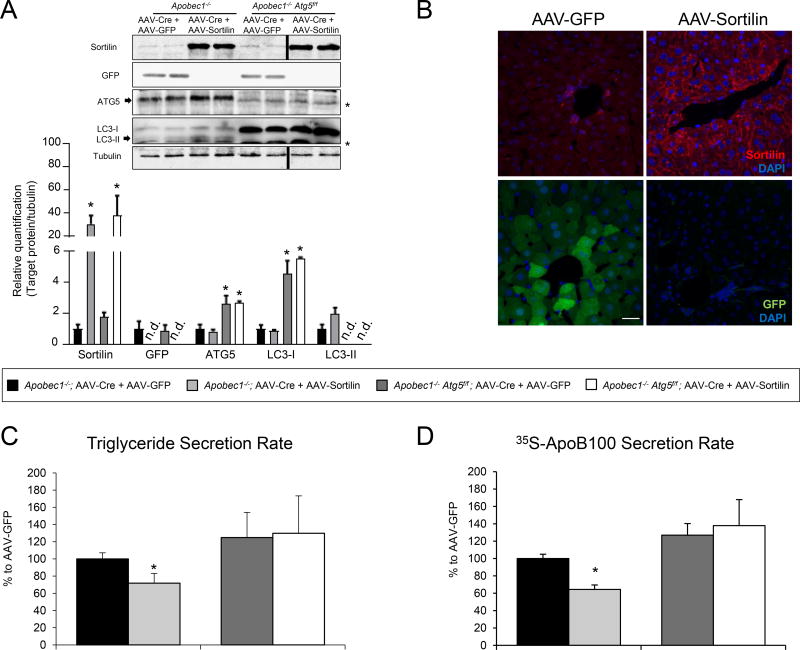
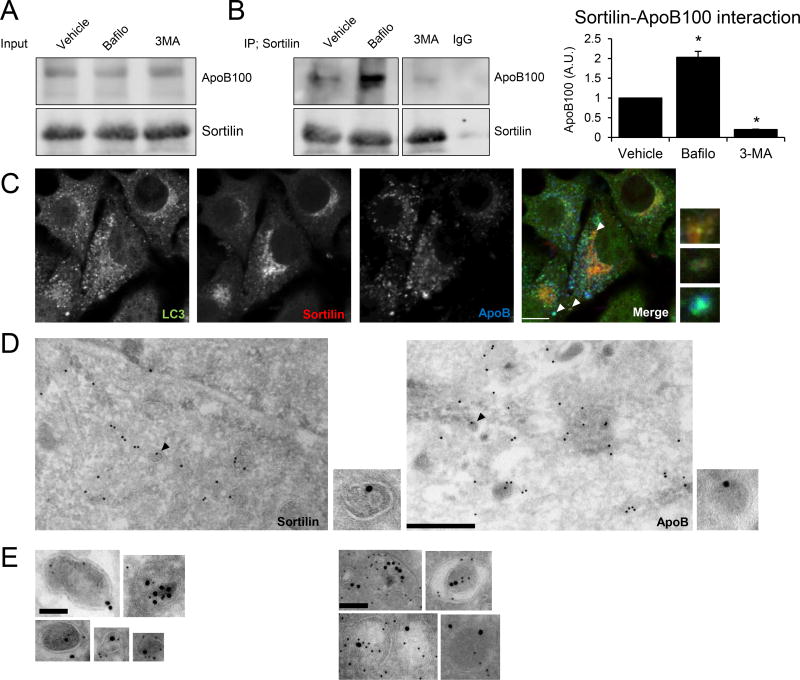
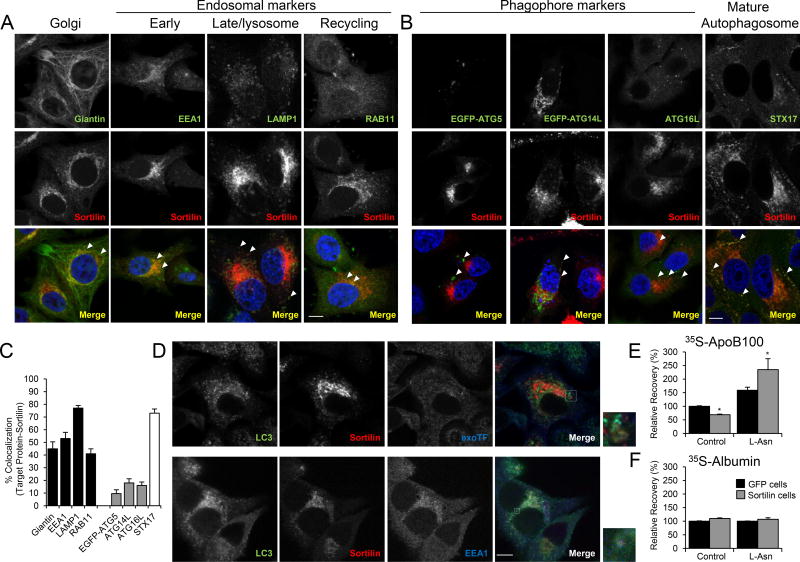
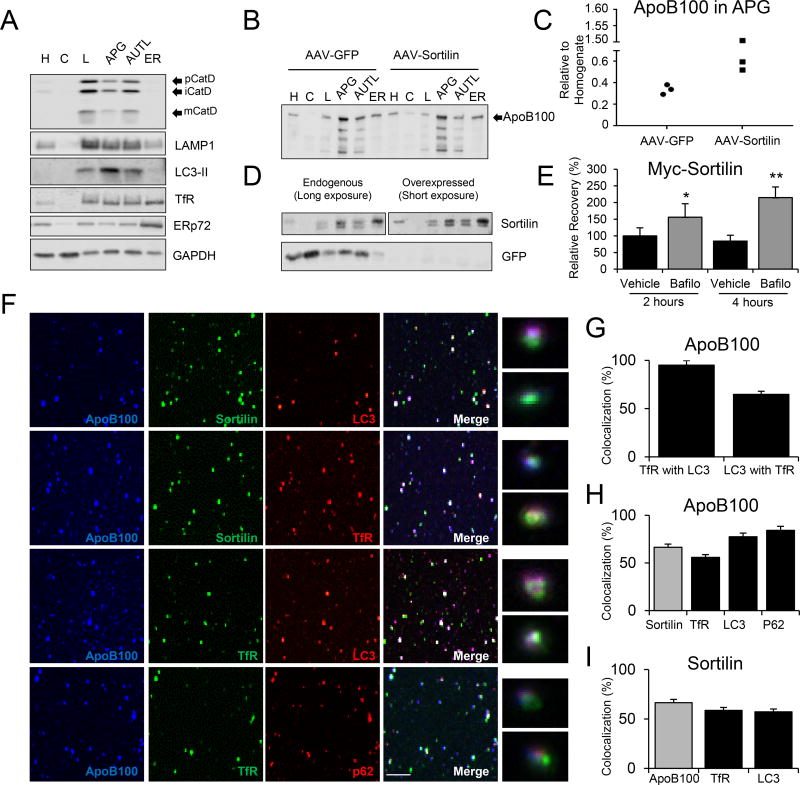
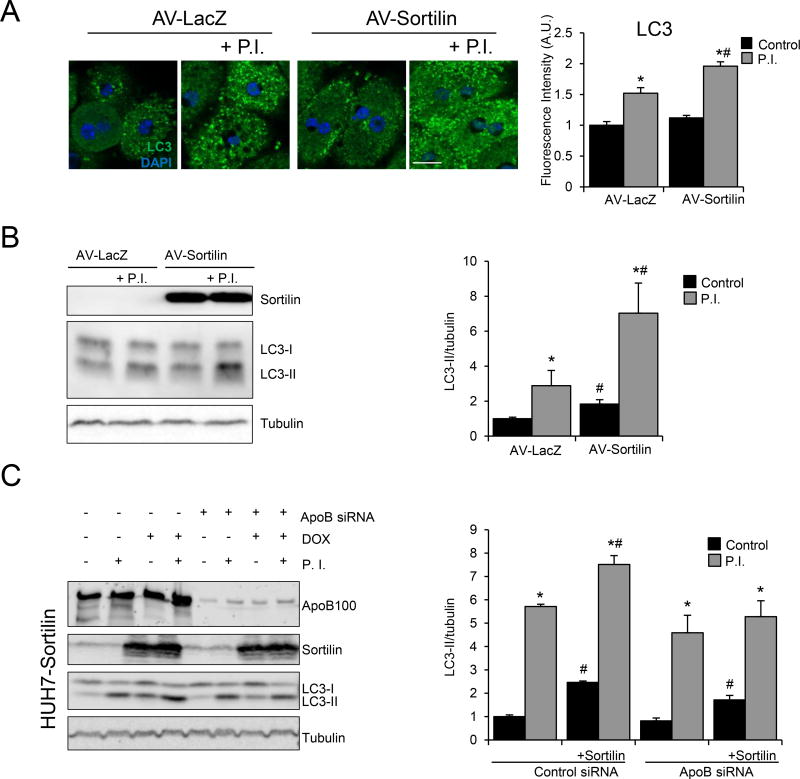
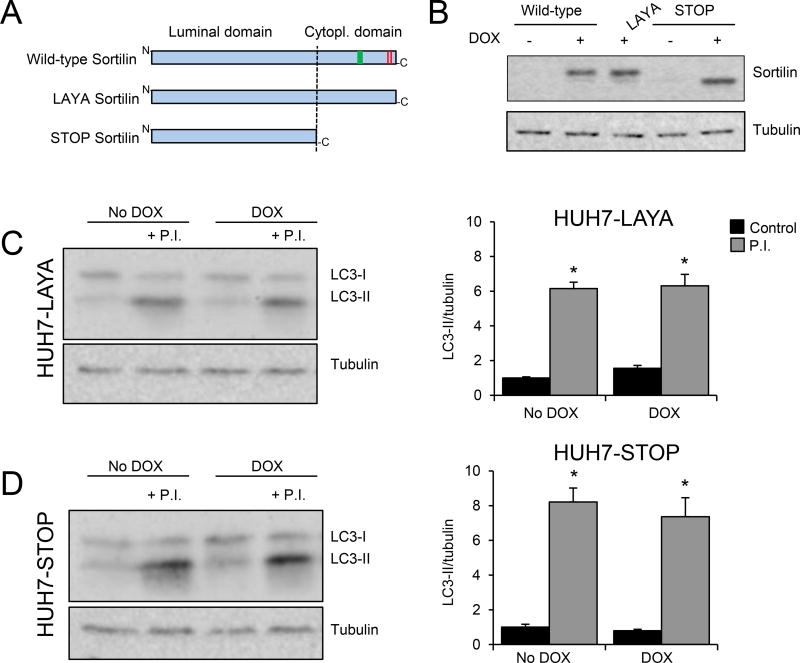
Methods and results: Pulse-chase studies directly established that SORT1 overexpression results in apoB degradation. As noted above, previous work implicated a role for lysosomes in this degradation. Through in vitro and in vivo studies, we now demonstrate that the sortilin-mediated route of apoB to lysosomes is unconventional and intersects with autophagy. Increased expression of sortilin diverts more apoB away from secretion, with both proteins trafficking to the endosomal compartment in vesicles that fuse with autophagosomes to form amphisomes. The amphisomes then merge with lysosomes. Furthermore, we show that sortilin itself is a regulator of autophagy and that its activity is scaled to the level of apoB synthesis.
Conclusions: These results strongly suggest that an unconventional lysosomal targeting process dependent on autophagy degrades apoB that was diverted from the secretory pathway by sortilin and provides a mechanism contributing to the reduced LDL-C found in individuals with SORT1 overexpression.
Keywords: amphisomes; apoB100; autophagy; liver; sortilin.
© 2018 American Heart Association, Inc.
Figures
References
-
- Teslovich TM, Musunuru K, Smith AV, Edmondson AC, Stylianou IM, Koseki M, Pirruccello JP, Ripatti S, Chasman DI, Willer CJ, Johansen CT, Fouchier SW, Isaacs A, Peloso GM, Barbalic M, et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature. 2010;466:707–13. - PMC - PubMed
-
- Musunuru K, Strong A, Frank-Kamenetsky M, Lee NE, Ahfeldt T, Sachs KV, Li X, Li H, Kuperwasser N, Ruda VM, Pirruccello JP, Muchmore B, Prokunina-Olsson L, Hall JL, Schadt EE, Morales CR, Lund-Katz S, Phillips MC, Wong J, Cantley W, Racie T, Ejebe KG, Orho-Melander M, Melander O, Koteliansky V, Fitzgerald K, Krauss RM, Cowan CA, Kathiresan S, Rader DJ. From noncoding variant to phenotype via SORT1 at the 1p13 cholesterol locus. Nature. 2010;466:714–9. - PMC - PubMed
-
- Calkhoven CF, Gringhuis SI, Ab G. The chicken CCAAT/enhancer binding protein alpha gene. Cloning, characterisation and tissue distribution. Gene. 1997;196:219–29. - PubMed
-
- Strong A, Ding Q, Edmondson AC, Millar JS, Sachs KV, Li X, Kumaravel A, Wang MY, Ai D, Guo L, Alexander ET, Nguyen D, Lund-Katz S, Phillips MC, Morales CR, Tall AR, Kathiresan S, Fisher EA, Musunuru K, Rader DJ. Hepatic sortilin regulates both apolipoprotein B secretion and LDL catabolism. The Journal of clinical investigation. 2012;122:2807–16. - PMC - PubMed
-
- Ai D, Baez JM, Jiang H, Conlon DM, Hernandez-Ono A, Frank-Kamenetsky M, Milstein S, Fitzgerald K, Murphy AJ, Woo CW, Strong A, Ginsberg HN, Tabas I, Rader DJ, Tall AR. Activation of ER stress and mTORC1 suppresses hepatic sortilin-1 levels in obese mice. The Journal of clinical investigation. 2012;122:1677–87. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
