

Deletion of a Long-Range Dlx5 Enhancer Disrupts Inner Ear Development in Mice
- PMID: 29301908
- PMCID: PMC5844329
- DOI: 10.1534/genetics.117.300447
Deletion of a Long-Range Dlx5 Enhancer Disrupts Inner Ear Development in Mice
Abstract
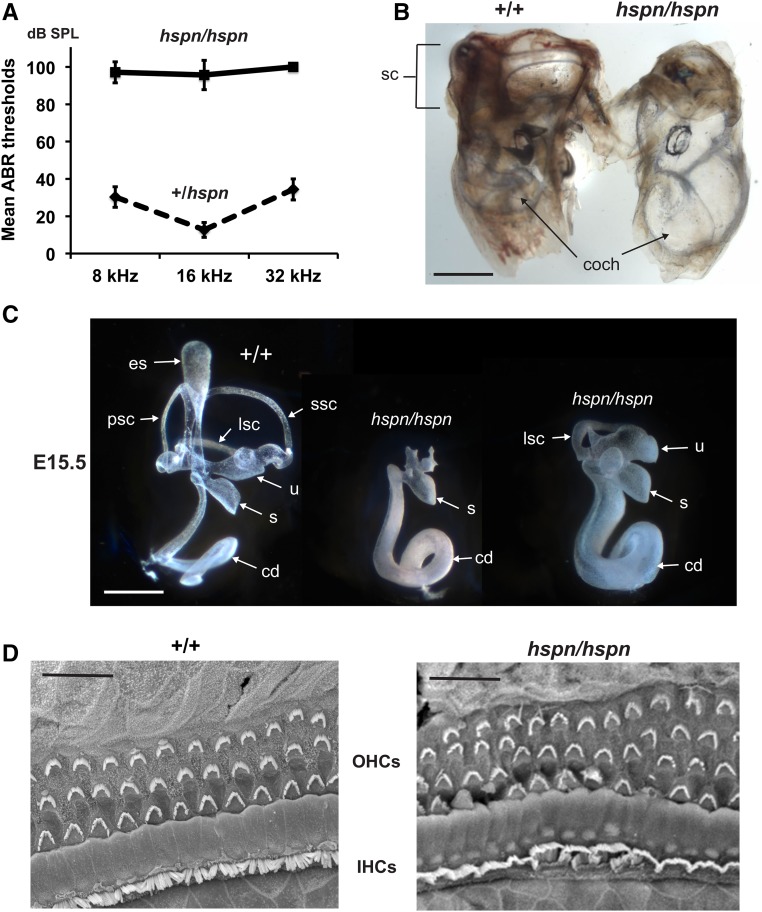
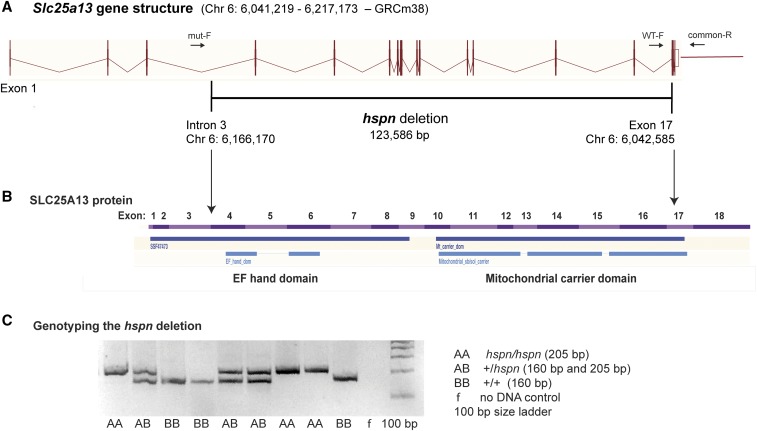
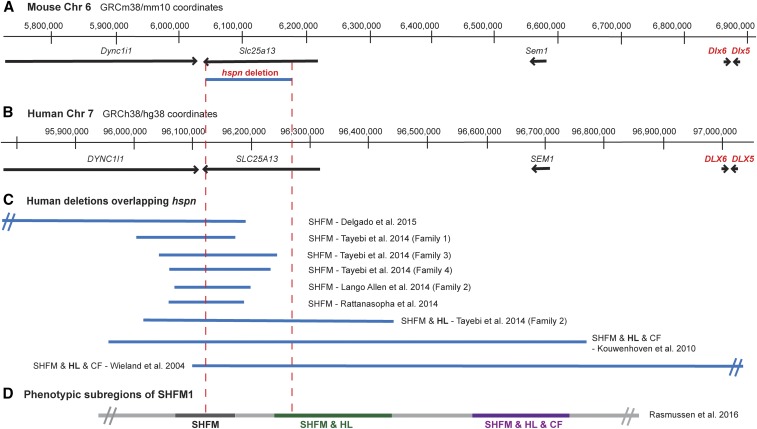
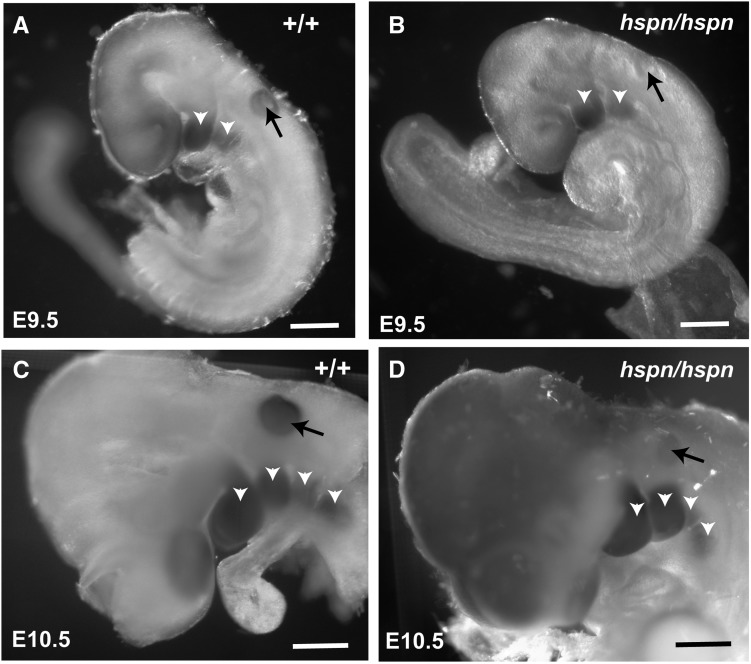
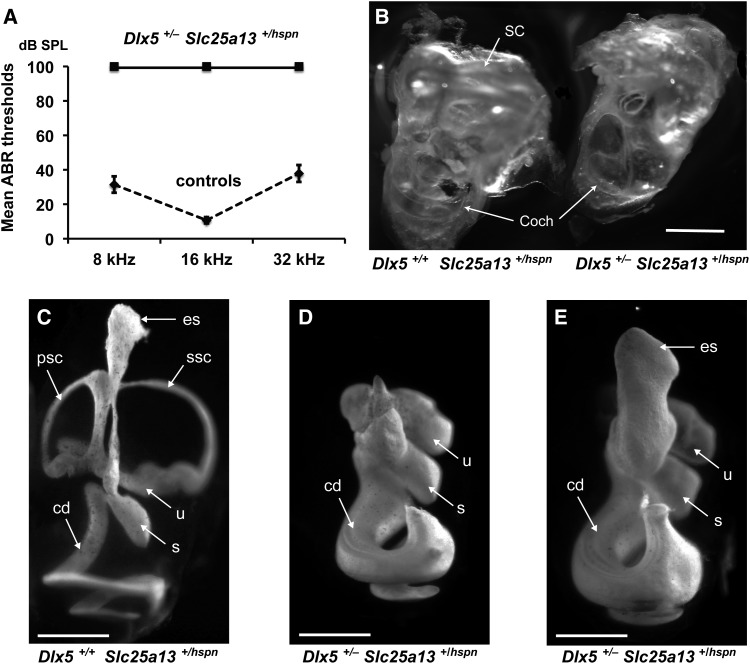
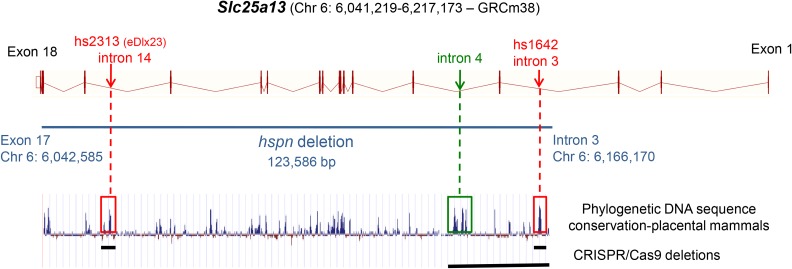
Distal enhancers are thought to play important roles in the spatiotemporal regulation of gene expression during embryonic development, but few predicted enhancer elements have been shown to affect transcription of their endogenous genes or to alter phenotypes when disrupted. Here, we demonstrate that a 123.6-kb deletion within the mouse Slc25a13 gene is associated with reduced transcription of Dlx5, a gene located 660 kb away. Mice homozygous for the Slc25a13 deletion mutation [named hyperspin (hspn)] have malformed inner ears and are deaf with balance defects, whereas previously reported Slc25a13 knockout mice showed no phenotypic abnormalities. Inner ears of Slc25a13hspn/hspn mice have malformations similar to those of Dlx5-/- embryos, and Dlx5 expression is severely reduced in the otocyst but not the branchial arches of Slc25a13hspn/hspn embryos, indicating that the Slc25a13hspn deletion affects otic-specific enhancers of Dlx5 In addition, transheterozygous Slc25a13+/hspn Dlx5+/- mice exhibit noncomplementation with inner ear dysmorphologies similar to those of Slc25a13hspn/hspn and Dlx5-/-embryos, verifying a cis-acting effect of the Slc25a13hspn deletion on Dlx5 expression. CRISPR/Cas9-mediated deletions of putative enhancer elements located within the Slc25a13hspn deleted region failed to phenocopy the defects of Slc25a13hspn/hspn mice, suggesting the possibility of multiple enhancers with redundant functions. Our findings in mice suggest that analogous enhancer elements in the human SLC25A13 gene may regulate DLX5 expression and underlie the hearing loss that is associated with split-hand/-foot malformation 1 syndrome. Slc25a13hspn/hspn mice provide a new animal model for studying long-range enhancer effects on Dlx5 expression in the developing inner ear.
Keywords: Dlx5; SHFM1; Slc25a13; distal enhancer; inner ear.
Copyright © 2018 by the Genetics Society of America.
Figures
Similar articles
-
Deletion of an enhancer near DLX5 and DLX6 in a family with hearing loss, craniofacial defects, and an inv(7)(q21.3q35).Hum Genet. 2010 Jan;127(1):19-31. doi: 10.1007/s00439-009-0736-4. Hum Genet. 2010. PMID: 19707792 Free PMC article.
-
Functional characterization of tissue-specific enhancers in the DLX5/6 locus.Hum Mol Genet. 2012 Nov 15;21(22):4930-8. doi: 10.1093/hmg/dds336. Epub 2012 Aug 21. Hum Mol Genet. 2012. PMID: 22914741 Free PMC article.
-
Identification of direct downstream targets of Dlx5 during early inner ear development.Hum Mol Genet. 2011 Apr 1;20(7):1262-73. doi: 10.1093/hmg/ddq567. Epub 2011 Jan 12. Hum Mol Genet. 2011. PMID: 21227998 Free PMC article.
-
Split Hand-Foot and Deafness in a Patient with 7q21.13-q21.3 Deletion Not Including the DLX5/6 Genes.Genes (Basel). 2023 Jul 26;14(8):1526. doi: 10.3390/genes14081526. Genes (Basel). 2023. PMID: 37628577 Free PMC article. Review.
-
Reassessing the Dlx code: the genetic regulation of branchial arch skeletal pattern and development.J Anat. 2005 Nov;207(5):501-61. doi: 10.1111/j.1469-7580.2005.00487.x. J Anat. 2005. PMID: 16313391 Free PMC article. Review.
Cited by
-
Potential of Gene and Cell Therapy for Inner Ear Hair Cells.Biomed Res Int. 2018 Jun 13;2018:8137614. doi: 10.1155/2018/8137614. eCollection 2018. Biomed Res Int. 2018. PMID: 30009175 Free PMC article. Review.
-
The noncoding genome and hearing loss.Hum Genet. 2022 Apr;141(3-4):323-333. doi: 10.1007/s00439-021-02359-z. Epub 2021 Sep 7. Hum Genet. 2022. PMID: 34491412 Review.
-
Deletion of a non-canonical regulatory sequence causes loss of Scn1a expression and epileptic phenotypes in mice.Genome Med. 2021 Apr 26;13(1):69. doi: 10.1186/s13073-021-00884-0. Genome Med. 2021. PMID: 33910599 Free PMC article.
-
Long-range cis-regulatory elements controlling GDF6 expression are essential for ear development.J Clin Invest. 2020 Aug 3;130(8):4213-4217. doi: 10.1172/JCI136951. J Clin Invest. 2020. PMID: 32369452 Free PMC article.
-
Interpreting the impact of noncoding structural variation in neurodevelopmental disorders.Genet Med. 2021 Jan;23(1):34-46. doi: 10.1038/s41436-020-00974-1. Epub 2020 Sep 25. Genet Med. 2021. PMID: 32973355 Free PMC article. Review.
References
-
- Acampora D., Merlo G. R., Paleari L., Zerega B., Postiglione M. P., et al. , 1999. Craniofacial, vestibular and bone defects in mice lacking the distal-less-related gene Dlx5. Development 126: 3795–3809. - PubMed
-
- Bernardini L., Palka C., Ceccarini C., Capalbo A., Bottillo I., et al. , 2008. Complex rearrangement of chromosomes 7q21.13-q22.1 confirms the ectrodactyly-deafness locus and suggests new candidate genes. Am. J. Med. Genet. A. 146A: 238–244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
