

Oxidation of the FAD cofactor to the 8-formyl-derivative in human electron-transferring flavoprotein
- PMID: 29301933
- PMCID: PMC5827430
- DOI: 10.1074/jbc.RA117.000846
Oxidation of the FAD cofactor to the 8-formyl-derivative in human electron-transferring flavoprotein
Abstract
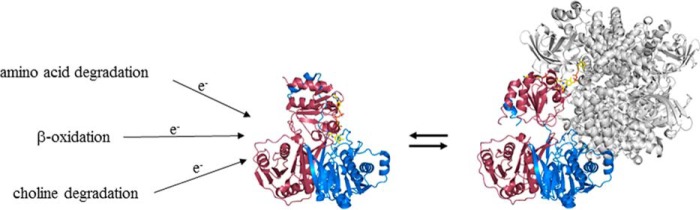
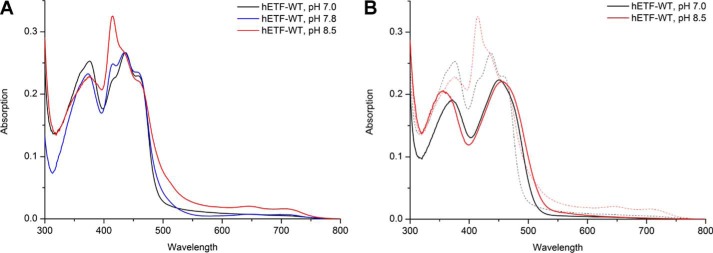
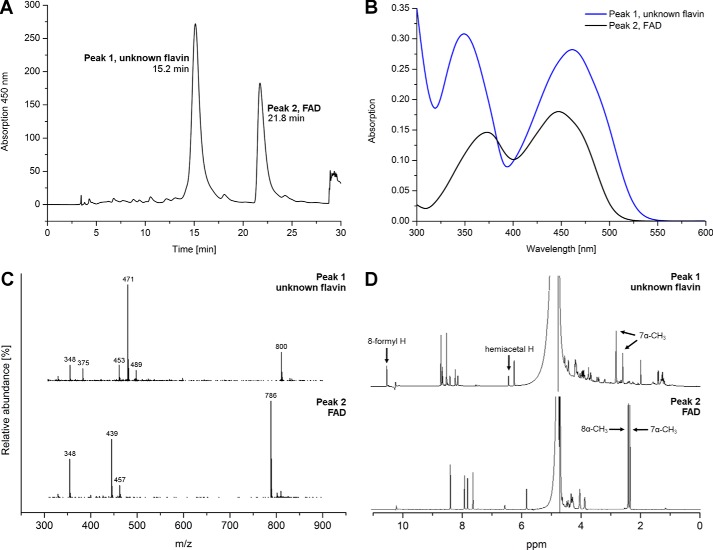
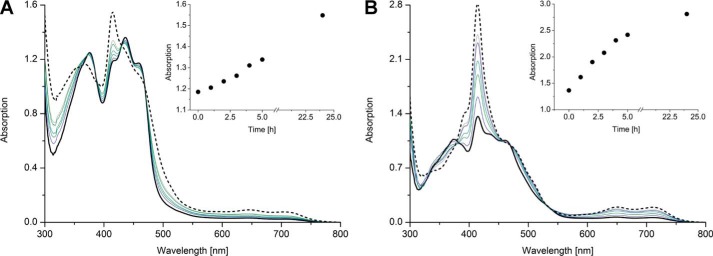
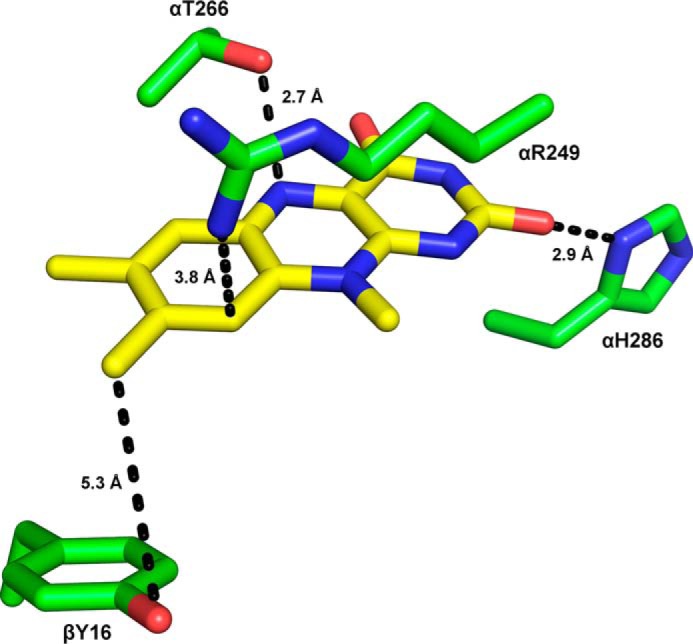
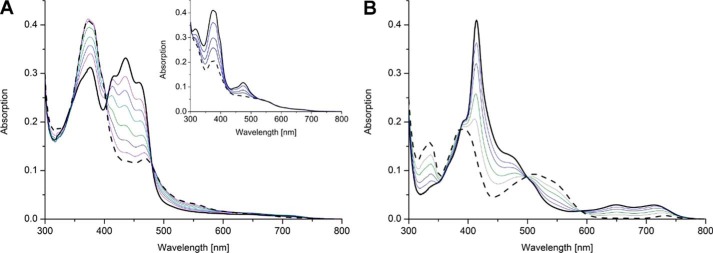
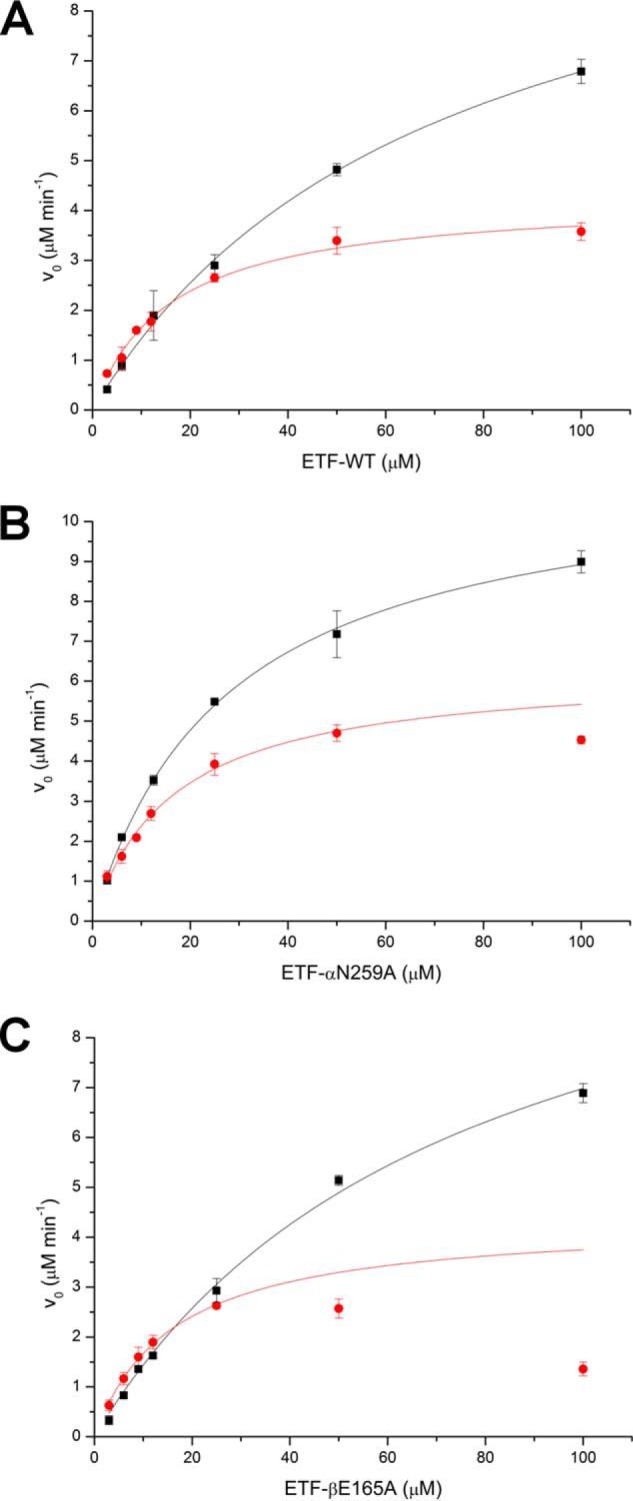
The heterodimeric human (h) electron-transferring flavoprotein (ETF) transfers electrons from at least 13 different flavin dehydrogenases to the mitochondrial respiratory chain through a non-covalently bound FAD cofactor. Here, we describe the discovery of an irreversible and pH-dependent oxidation of the 8α-methyl group to 8-formyl-FAD (8f-FAD), which represents a unique chemical modification of a flavin cofactor in the human flavoproteome. Furthermore, a set of hETF variants revealed that several conserved amino acid residues in the FAD-binding pocket of electron-transferring flavoproteins are required for the conversion to the formyl group. Two of the variants generated in our study, namely αR249C and αT266M, cause glutaric aciduria type II, a severe inherited disease. Both of the variants showed impaired formation of 8f-FAD shedding new light on the potential molecular cause of disease development. Interestingly, the conversion of FAD to 8f-FAD yields a very stable flavin semiquinone that exhibited slightly lower rates of electron transfer in an artificial assay system than hETF containing FAD. In contrast, the formation of 8f-FAD enhanced the affinity to human dimethylglycine dehydrogenase 5-fold, indicating that formation of 8f-FAD modulates the interaction of hETF with client enzymes in the mitochondrial matrix. Thus, we hypothesize that the FAD cofactor bound to hETF is subject to oxidation in the alkaline (pH 8) environment of the mitochondrial matrix, which may modulate electron transport between client dehydrogenases and the respiratory chain. This discovery challenges the current concepts of electron transfer processes in mitochondria.
Keywords: 8-formyl-FAD; dehydrogenase; electron transfer; flavin adenine dinucleotide (FAD); flavin semiquinone; mitochondria; respiratory chain.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
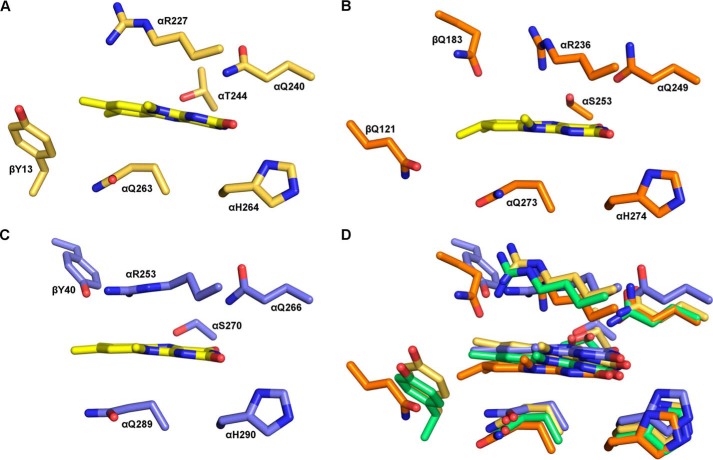
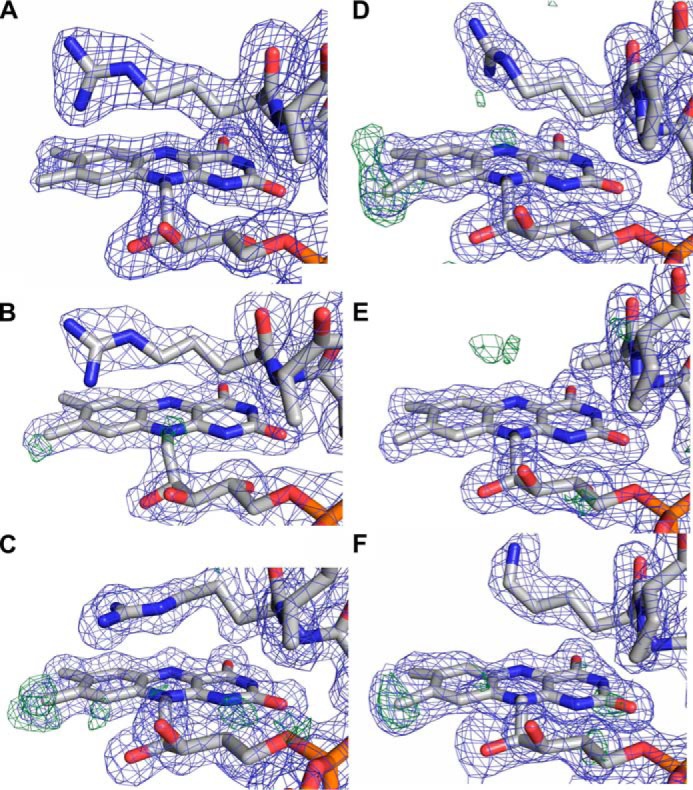
References
-
- Crane F. L., Mii S., Hauge J. G., Green D. E., and Beinert H. (1956) On the mechanism of dehydrogenation of fatty acyl derivatives of coenzyme A. I. The general fatty acyl coenzyme A dehydrogenase. J. Biol. Chem. 218, 701–706 - PubMed
-
- Toogood H. S., van Thiel A., Basran J., Sutcliffe M. J., Scrutton N. S., and Leys D. (2004) Extensive domain motion and electron transfer in the human electron-transferring flavoprotein·medium chain acyl-CoA dehydrogenase complex. J. Biol. Chem. 279, 32904–32912 10.1074/jbc.M404884200 - DOI - PubMed
-
- Herrick K. R., Salazar D., Goodman S. I., Finocchiaro G., Bedzyk L. A., and Frerman F. E. (1994) Expression and characterization of human and chimeric human-Paracoccus denitrificans electron transfer flavoproteins. J. Biol. Chem. 269, 32239–32245 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
