

Invading, Leading and Navigating Cells in Caenorhabditis elegans: Insights into Cell Movement in Vivo
- PMID: 29301948
- PMCID: PMC5753875
- DOI: 10.1534/genetics.117.300082
Invading, Leading and Navigating Cells in Caenorhabditis elegans: Insights into Cell Movement in Vivo
Abstract
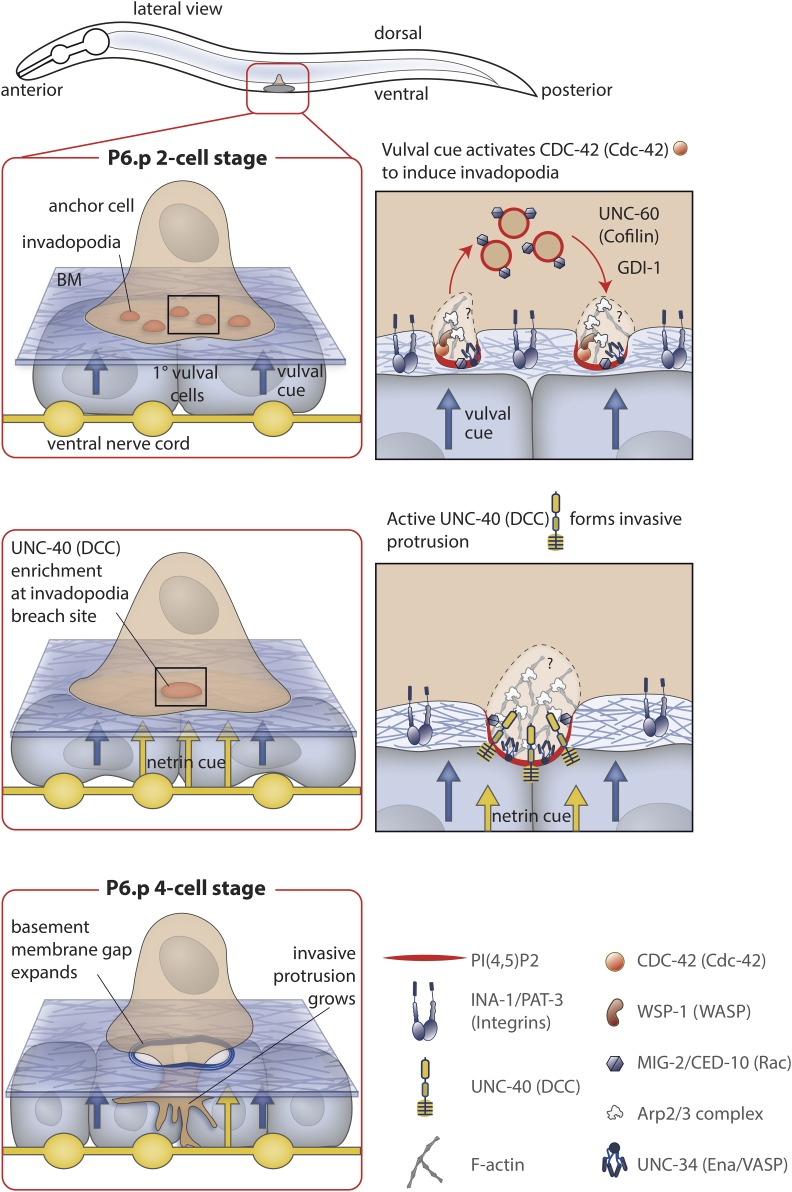
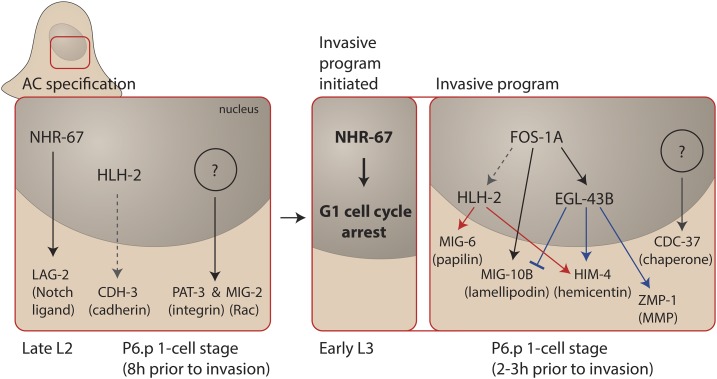
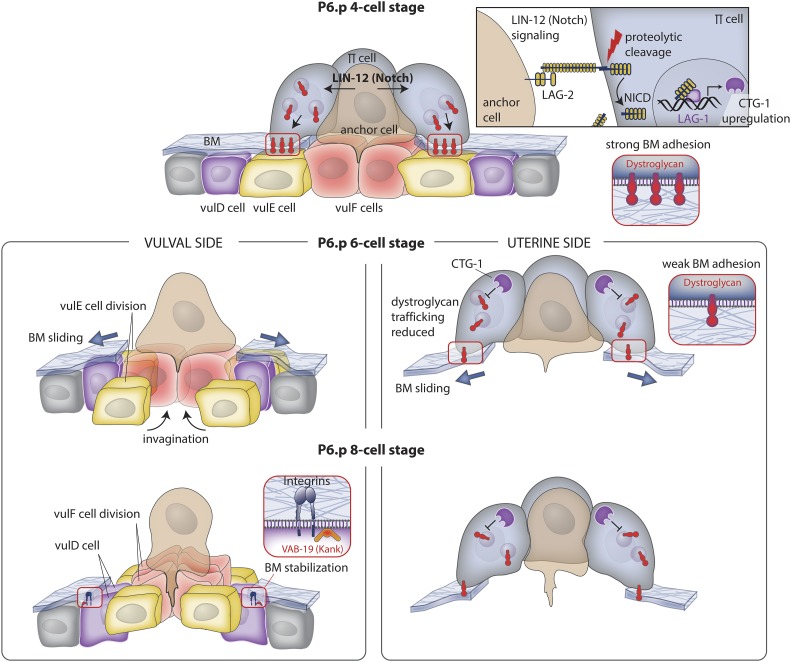
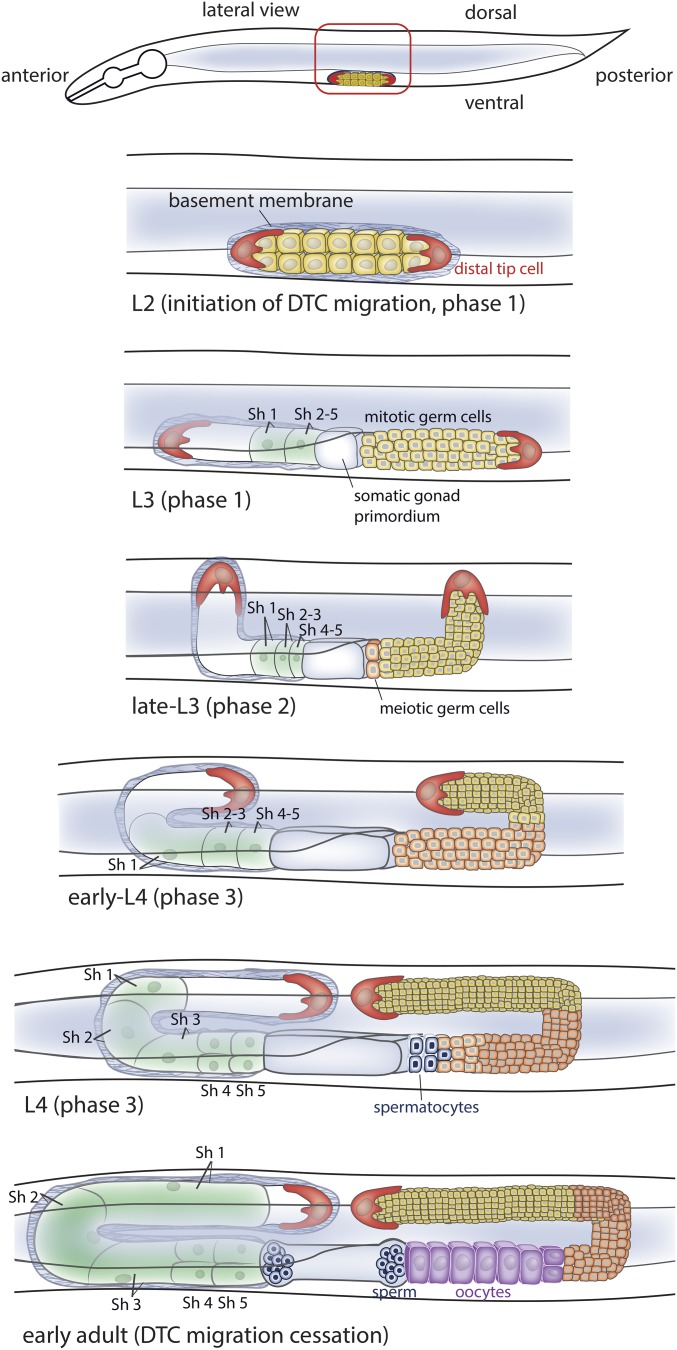
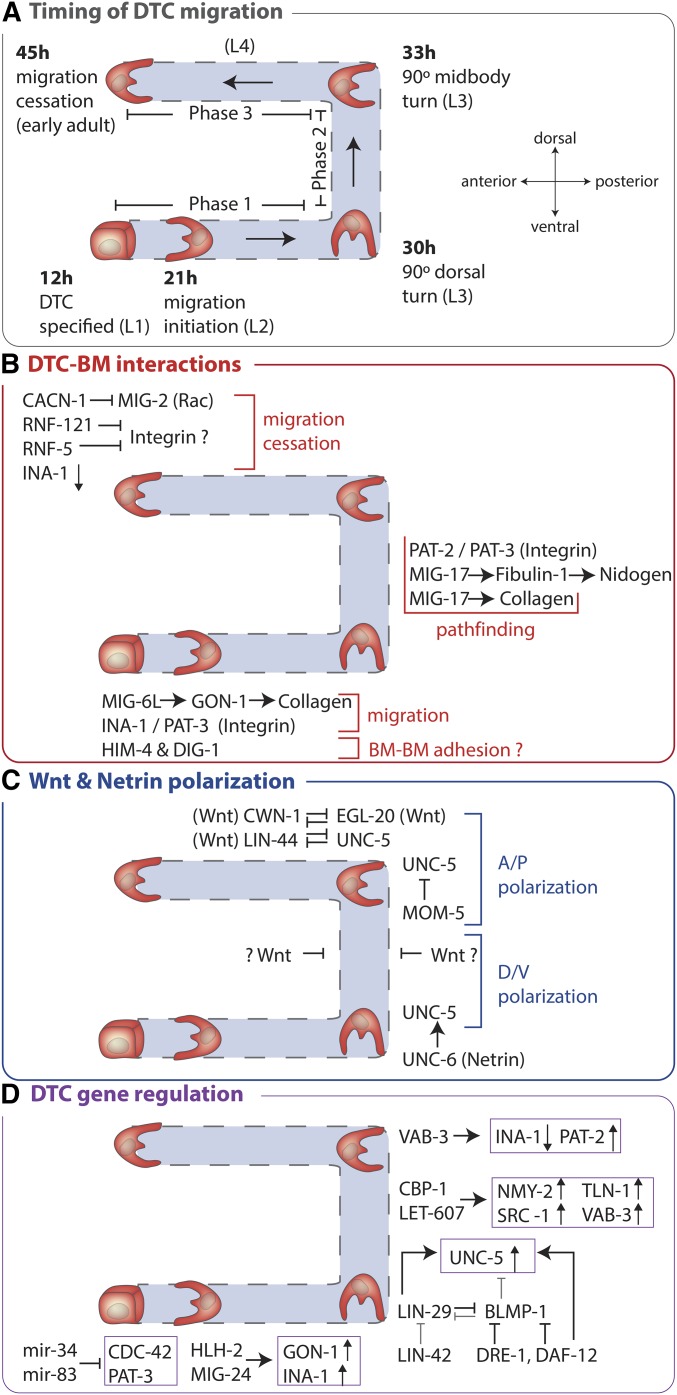
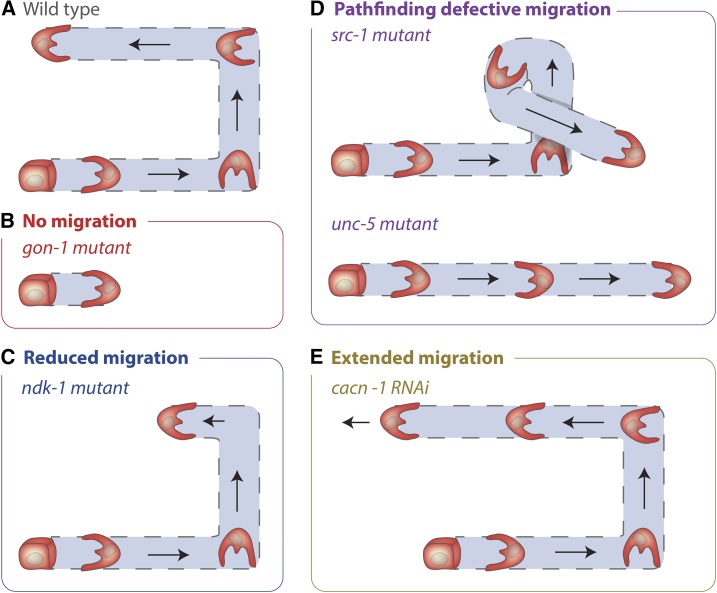
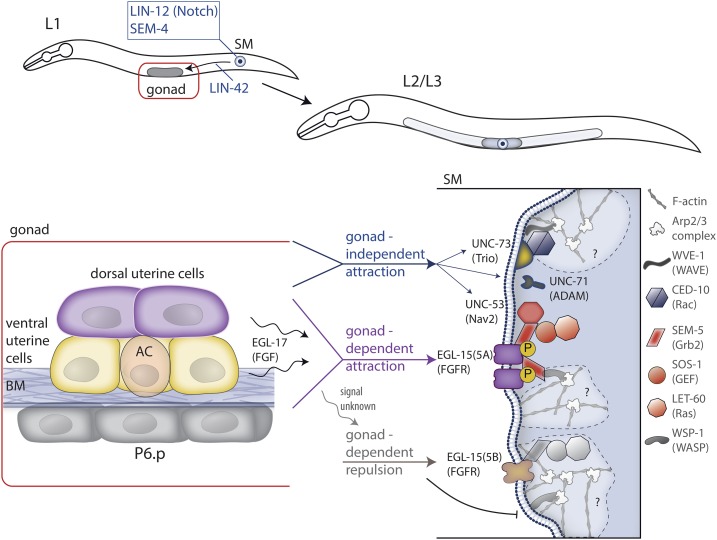
Highly regulated cell migration events are crucial during animal tissue formation and the trafficking of cells to sites of infection and injury. Misregulation of cell movement underlies numerous human diseases, including cancer. Although originally studied primarily in two-dimensional in vitro assays, most cell migrations in vivo occur in complex three-dimensional tissue environments that are difficult to recapitulate in cell culture or ex vivo Further, it is now known that cells can mobilize a diverse repertoire of migration modes and subcellular structures to move through and around tissues. This review provides an overview of three distinct cellular movement events in Caenorhabditis elegans-cell invasion through basement membrane, leader cell migration during organ formation, and individual cell migration around tissues-which together illustrate powerful experimental models of diverse modes of movement in vivo We discuss new insights into migration that are emerging from these in vivo studies and important future directions toward understanding the remarkable and assorted ways that cells move in animals.
Keywords: C. elegans; F-actin; FGF pathway; Wnt pathway; WormBook; basement membrane; cell invasion; cell migration; cell signaling; integrin; netrin pathway.
Copyright © 2018 by the Genetics Society of America.
Figures
References
-
- Abella J. V., Galloni C., Pernier J., Barry D. J., Kjaer S., et al. , 2016. Isoform diversity in the Arp2/3 complex determines actin filament dynamics. Nat. Cell Biol. 18: 76–86. - PubMed
-
- Altincicek B., Fischer M., Luersen K., Boll M., Wenzel U., et al. , 2010. Role of matrix metalloproteinase ZMP-2 in pathogen resistance and development in Caenorhabditis elegans. Dev. Comp. Immunol. 34: 1160–1169. - PubMed
-
- Aman A., Piotrowski T., 2010. Cell migration during morphogenesis. Dev. Biol. 341: 20–33. - PubMed
-
- Antebi A., Culotti J. G., Hedgecock E. M., 1998. daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development 125: 1191–1205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
