

Cell-Cycle-Specific Function of p53 in Fanconi Anemia Hematopoietic Stem and Progenitor Cell Proliferation
- PMID: 29307578
- PMCID: PMC5830889
- DOI: 10.1016/j.stemcr.2017.12.006
Cell-Cycle-Specific Function of p53 in Fanconi Anemia Hematopoietic Stem and Progenitor Cell Proliferation
Abstract
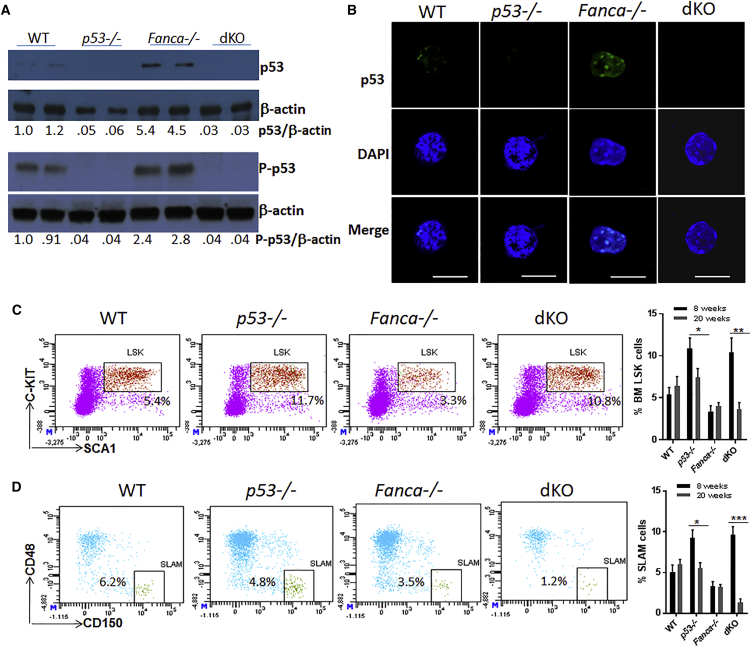
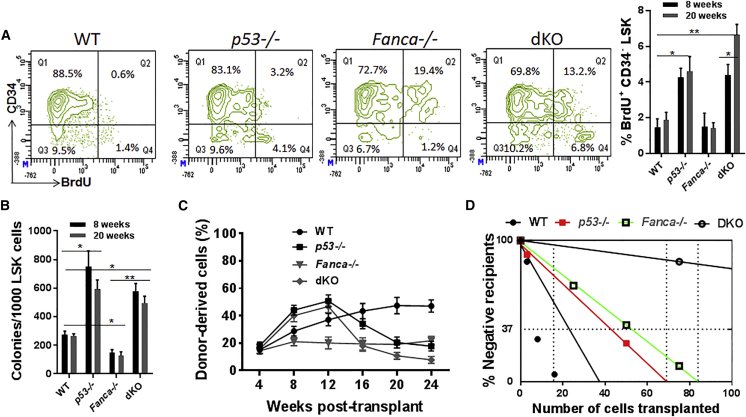
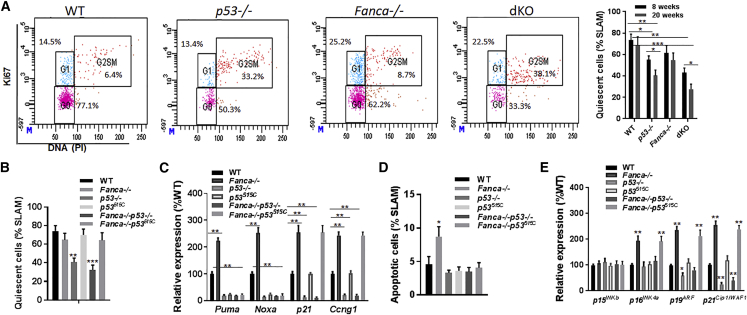
Overactive p53 has been proposed as an important pathophysiological factor for bone marrow failure syndromes, including Fanconi anemia (FA). Here, we report a p53-dependent effect on hematopoietic stem and progenitor cell (HSPC) proliferation in mice deficient for the FA gene Fanca. Deletion of p53 in Fanca-/- mice leads to replicative exhaustion of the hematopoietic stem cell (HSC) in transplant recipients. Using Fanca-/- HSCs expressing the separation-of-function mutant p53515C transgene, which selectively impairs the p53 function in apoptosis but keeps its cell-cycle checkpoint activities intact, we show that the p53 cell-cycle function is specifically required for the regulation of Fanca-/- HSC proliferation. Our results demonstrate that p53 plays a compensatory role in preventing FA HSCs from replicative exhaustion and suggest a cautious approach to manipulating p53 signaling as a therapeutic utility in FA.
Keywords: Fanconi anemia; apoptosis; bone marrow failure; cell cycle; hematopoietic stem and progenitor cells; p53; proliferation.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
