

An Ichor-dependent apical extracellular matrix regulates seamless tube shape and integrity
- PMID: 29309404
- PMCID: PMC5774827
- DOI: 10.1371/journal.pgen.1007146
An Ichor-dependent apical extracellular matrix regulates seamless tube shape and integrity
Abstract
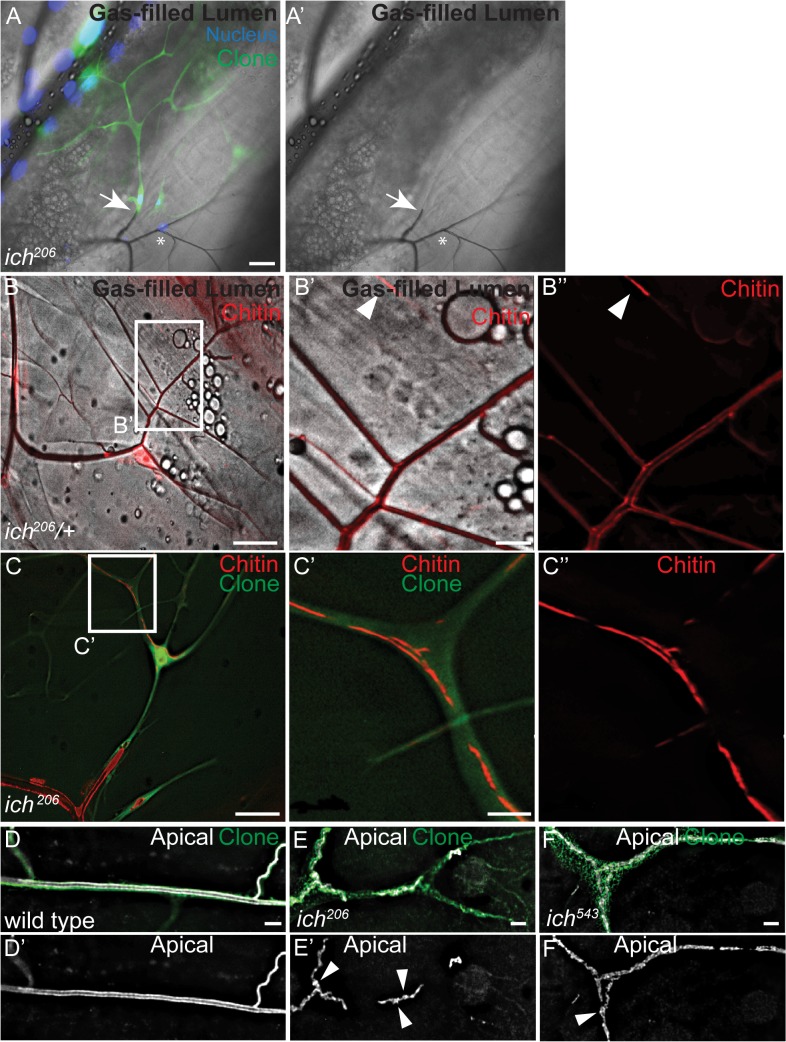
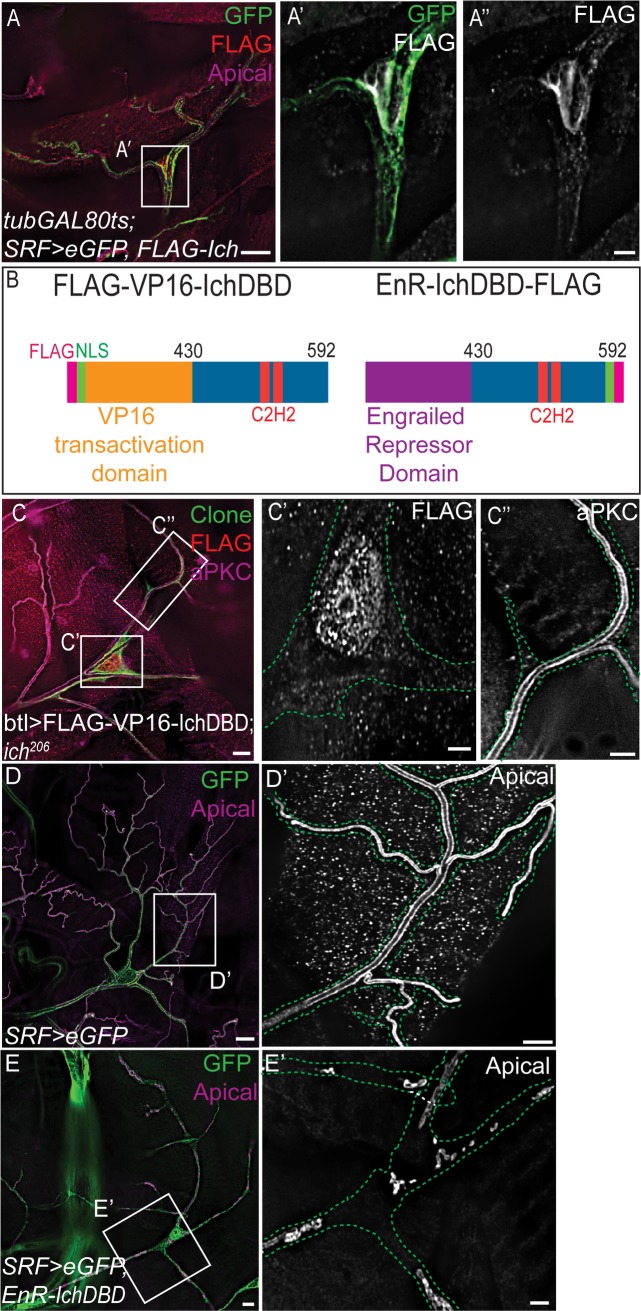
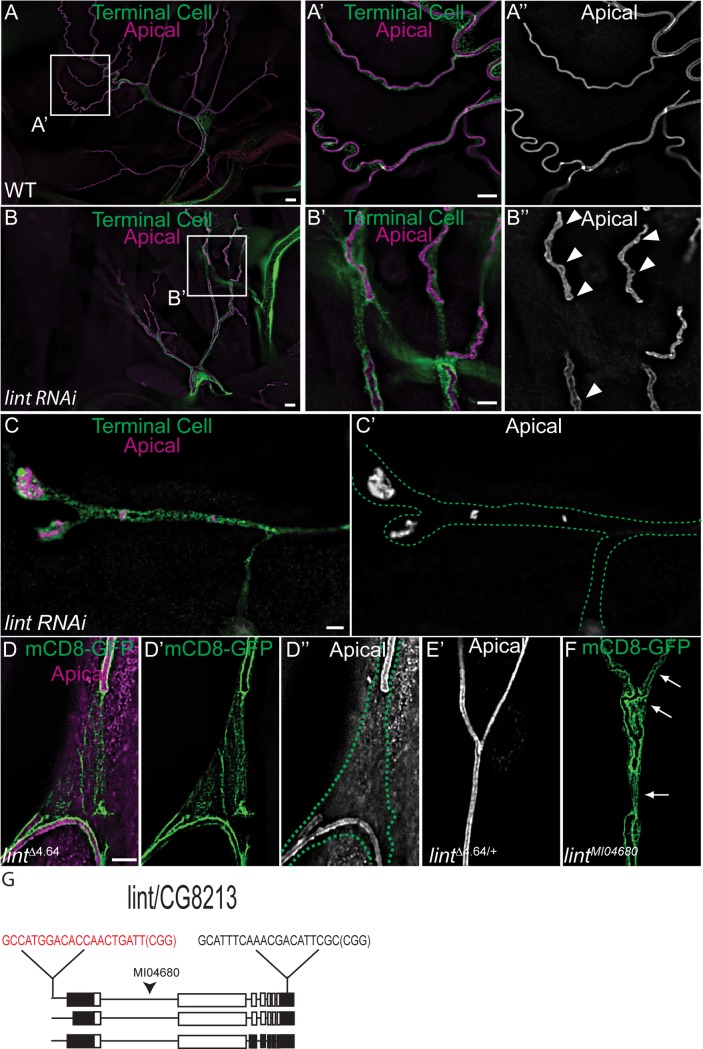
During sprouting angiogenesis in the vertebrate vascular system, and primary branching in the Drosophila tracheal system, specialized tip cells direct branch outgrowth and network formation. When tip cells lumenize, they form subcellular (seamless) tubes. How these seamless tubes are made, shaped and maintained remains poorly understood. Here we characterize a Drosophila mutant called ichor (ich), and show that ich is essential for the integrity and shape of seamless tubes in tracheal terminal cells. We find that Ich regulates seamless tubulogenesis via its role in promoting the formation of a mature apical extracellular matrix (aECM) lining the lumen of the seamless tubes. We determined that ich encodes a zinc finger protein (CG11966) that acts, as a transcriptional activator required for the expression of multiple aECM factors, including a novel membrane-anchored trypsin protease (CG8213). Thus, the integrity and shape of seamless tubes are regulated by the aECM that lines their lumens.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Fedeles SV, Tian X, Gallagher A-R, Mitobe M, Nishio S, Lee SH, et al. A genetic interaction network of five genes for human polycystic kidney and liver diseases defines polycystin-1 as the central determinant of cyst formation. Nature Genetics. 2011. June 19;43(7):639–47. doi: 10.1038/ng.860 - DOI - PMC - PubMed
-
- Tual-Chalot S, Oh SP, Arthur HM. Mouse models of hereditary hemorrhagic telangiectasia: recent advances and future challenges. Frontiers in Genetics [Internet]. 2015. February 18; 6 Available from: http://journal.frontiersin.org/Article/10.3389/fgene.2015.00025/abstract. - DOI - PMC - PubMed
-
- Lubarsky B, Krasnow MA. Tube Morphogenesis: making and shaping biological tubes. Cell. 2003. January;112(1):19–28. - PubMed
-
- Buechner M. Tubes and the single C. elegans excretory cell. Trends in Cell Biology. 2002. October 1;12(10):479–84. - PubMed
-
- Berry KL. A C. elegans CLIC-like Protein Required for Intracellular Tube Formation and Maintenance. Science. 2003. December 19;302(5653):2134–7. doi: 10.1126/science.1087667 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
