

Co-expression of a cyclizing asparaginyl endopeptidase enables efficient production of cyclic peptides in planta
- PMID: 29309615
- PMCID: PMC5853369
- DOI: 10.1093/jxb/erx422
Co-expression of a cyclizing asparaginyl endopeptidase enables efficient production of cyclic peptides in planta
Abstract
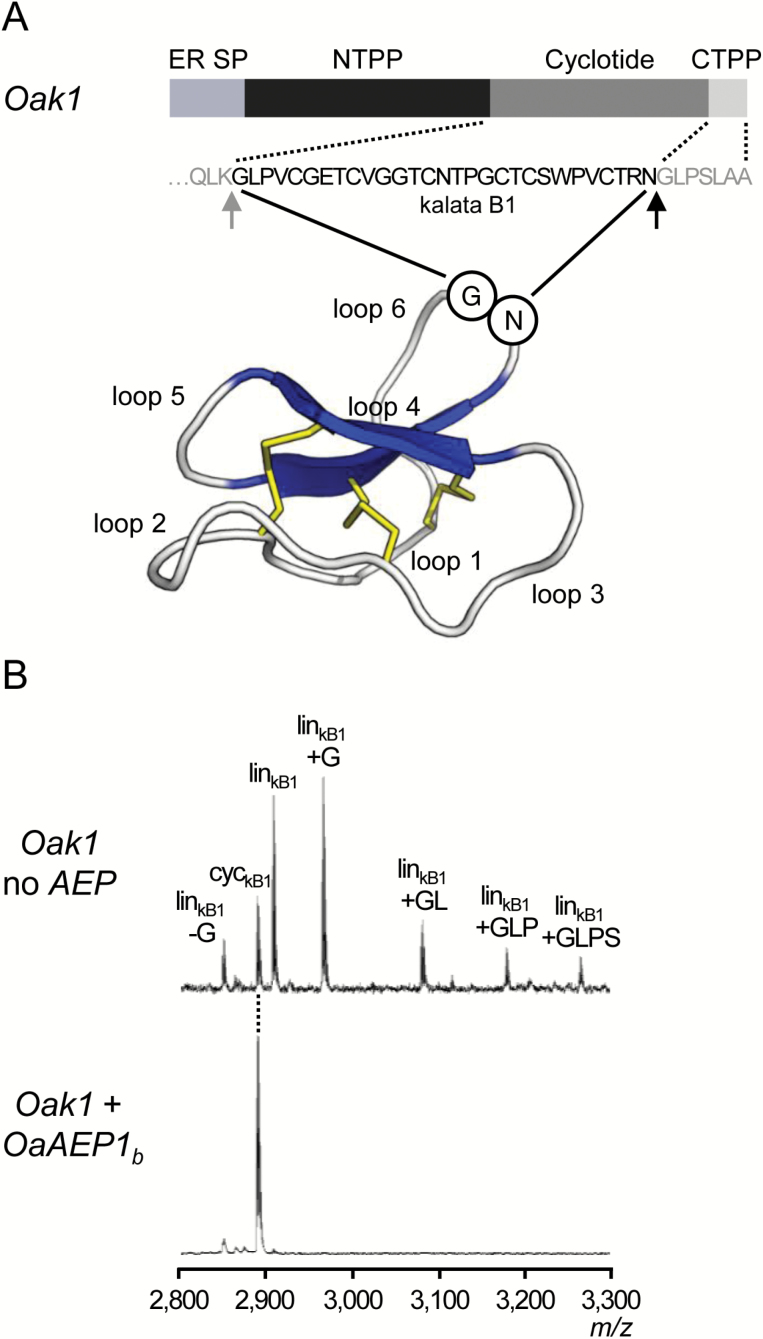
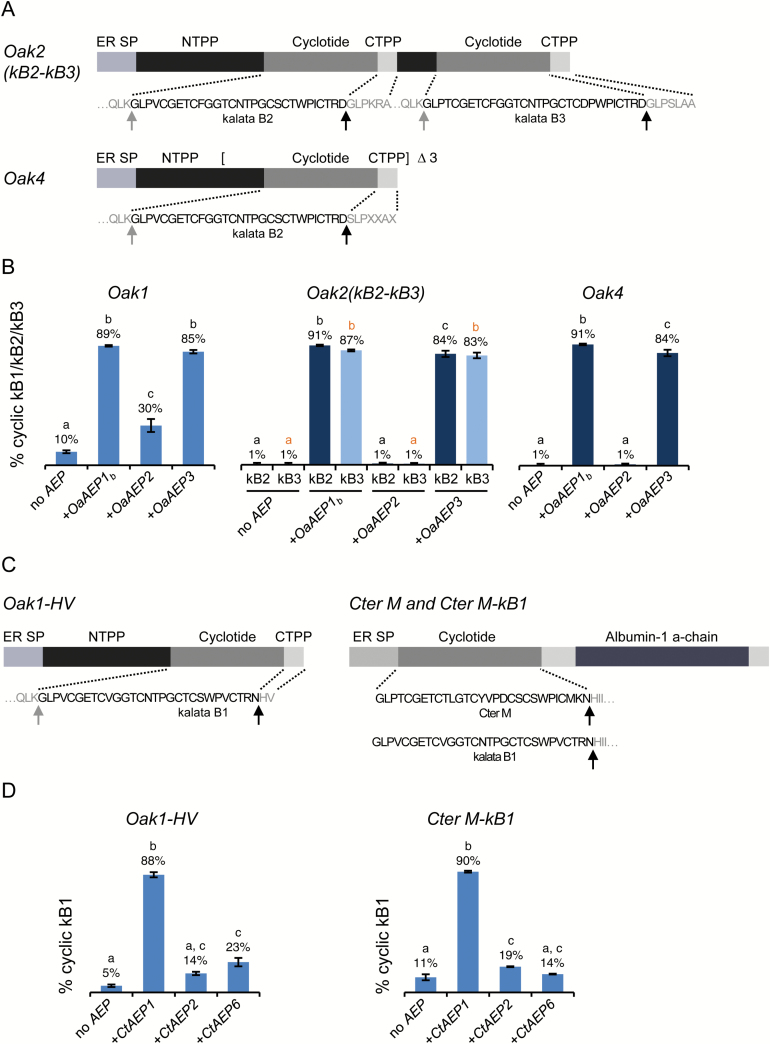
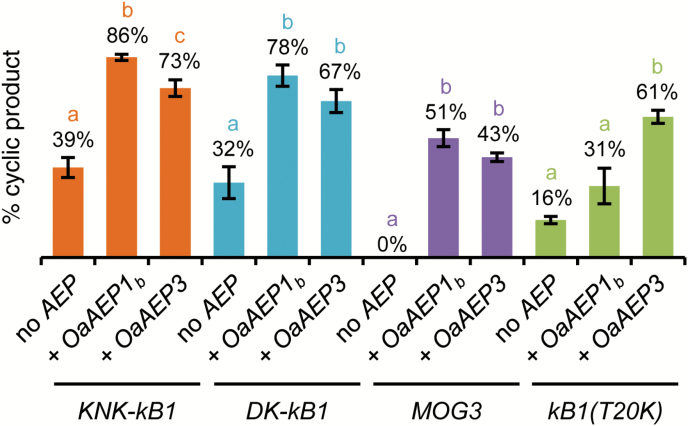
Cyclotides are ultra-stable, backbone-cyclized plant defence peptides that have attracted considerable interest in the pharmaceutical industry. This is due to their range of native bioactivities as well as their ability to stabilize other bioactive peptides within their framework. However, a hindrance to their widespread application is the lack of scalable, cost-effective production strategies. Plant-based production is an attractive, benign option since all biosynthetic steps are performed in planta. Nonetheless, cyclization in non-cyclotide-producing plants is poor. Here, we show that cyclic peptides can be produced efficiently in Nicotiana benthamiana, one of the leading plant-based protein production platforms, by co-expressing cyclotide precursors with asparaginyl endopeptidases that catalyse peptide backbone cyclization. This approach was successful in a range of other plants (tobacco, bush bean, lettuce, and canola), either transiently or stably expressed, and was applicable to both native and engineered cyclic peptides. We also describe the use of the transgenic system to rapidly identify new asparaginyl endopeptidase cyclases and interrogate their substrate sequence requirements. Our results pave the way for exploiting cyclotides for pest protection in transgenic crops as well as large-scale production of cyclic peptide pharmaceuticals in plants.
Keywords: Asparaginyl endopeptidase; Nicotiana benthamiana; SFTI; cyclic peptide; cyclotide; kalata B1; plant-made pharmaceutical; transient expression.
© The Author(s) 2017. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
