

doi: 10.1182/blood-2017-09-808402.
Epub 2018 Jan 8.
Five percent of healthy newborns have an ETV6-RUNX1 fusion as revealed by DNA-based GIPFEL screening
Affiliations
- PMID: 29311095
- PMCID: PMC5909885
- DOI: 10.1182/blood-2017-09-808402
Item in Clipboard
Five percent of healthy newborns have an ETV6-RUNX1 fusion as revealed by DNA-based GIPFEL screening
Blood.
.
No abstract available
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
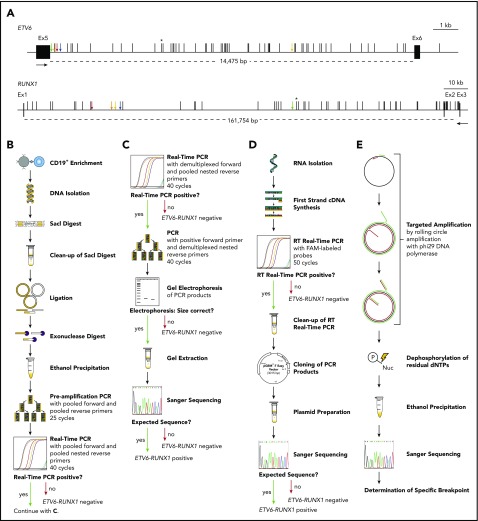
The incidence of ETV6-RUNX1 fusions in healthy newborns can be determined by DNA-based GIPFEL screening. (A) Only DNA- and not RNA-based ETV6-RUNX1 detection provides individual-specific breakpoints. The breakpoint cluster regions (BCRs; horizontal lines, exact regions are marked by dotted lines, sizes are given in base pairs) of ETV6 (top panel, intron 5) and RUNX1 (bottom panel, intron 1 and 2) are presented. Black boxes indicate exons and vertical black lines on top of the BCRs indicate reported patient-specific breakpoints.- Asterisks mark the breakpoint present in the REH cell line. The breakpoints identified in the present study are indicated by orange (N424), blue (N726), red (N817), green (N823), and yellow (N890) arrows. Black arrows represent primers that are usually used for ETV6-RUNX1 screening by RT-PCR. They generate 1 of 2 PCR products for every possible breakpoint within the BCRs: exon 5 of ETV6 fused to either exon 2 or exon 3 of RUNX1. Only DNA-based techniques can differentiate between breakpoints that are specific for each individual patient and localized in intronic regions. (B-E) The workflow of cord blood screening using a modified GIPFEL technique is presented. (B) CD19+ B cells were enriched from newborn cord blood. DNA was isolated, fragmented using SacI, and purified. The DNA was ligated to achieve circularization and residual linear DNA was digested. DNA circles were purified and PCR (preamplification) and real-time PCR were used to detect the ligation joints produced by circularization. The protocol was carried out with each sample. (C) Optional continuation protocol that was carried out if the real-time PCR in panel B generated a putative positive result. Then a new real-time PCR was carried out with demultiplexed forward primers. If 1 of the forward primers produced a positive result, the PCR was repeated with this forward primer and demultiplexed reverse primers. The products were then analyzed on an agarose gel and Sanger sequenced. In case of negative results, no further validation steps were done. (D) Workflow of the transcriptional validation. RNA was reverse transcribed and subjected to real-time PCR. In case of a putative positive result, the PCR product was purified, cloned, and subjected to Sanger sequencing. (E) Further development of the original GIPFEL technique allowed the identification of patient-specific breakpoints with base-pair resolution. To this end, circularized DNA was amplified using specific primers that hybridized to the known ligation joint region and faced toward the unknown breakpoint. Amplification was carried out with Phi29 DNA polymerase, leading to linear products. With Phi29, amplification occurs at a constant temperature and new primers can constantly bind to the original and the amplified DNA, leading to further amplification. The amplified DNA was subsequently purified and Sanger sequenced. cDNA, complementary DNA; dNTP, deoxynucleotide triphosphate; Ex, exon; FAM, fluorescein amidite; Nuc, deoxynucleoside diphospate; P, phosphate.
References
-
- Wiemels JL, Ford AM, Van Wering ER, Postma A, Greaves M. Protracted and variable latency of acute lymphoblastic leukemia after TEL-AML1 gene fusion in utero. Blood. 1999;94(3):1057-1062. - PubMed
-
- Eguchi-Ishimae M, Eguchi M, Ishii E, et al. Breakage and fusion of the TEL (ETV6) gene in immature B lymphocytes induced by apoptogenic signals. Blood. 2001;97(3):737-743. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
