

Dynamic changes in carbonate chemistry in the microenvironment around single marine phytoplankton cells
- PMID: 29311545
- PMCID: PMC5758611
- DOI: 10.1038/s41467-017-02426-y
Dynamic changes in carbonate chemistry in the microenvironment around single marine phytoplankton cells
Abstract
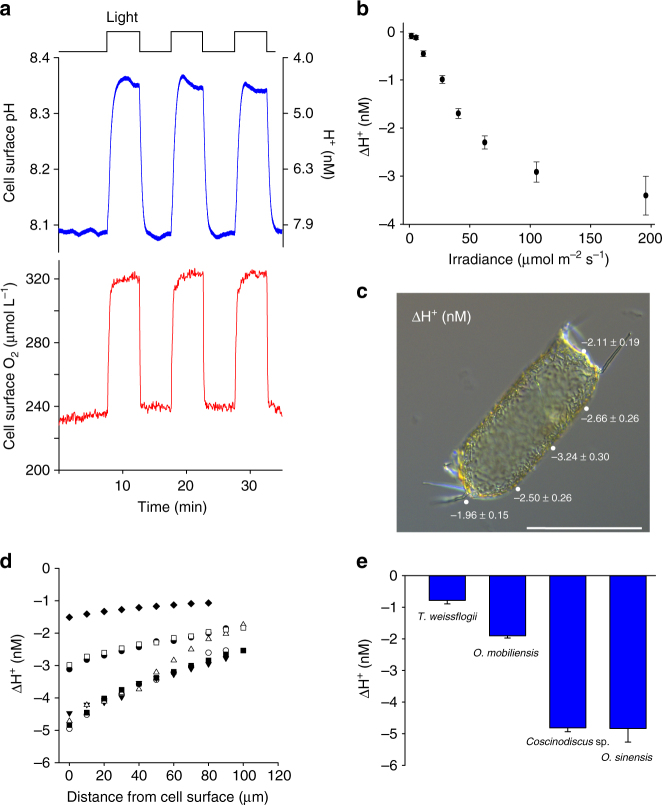
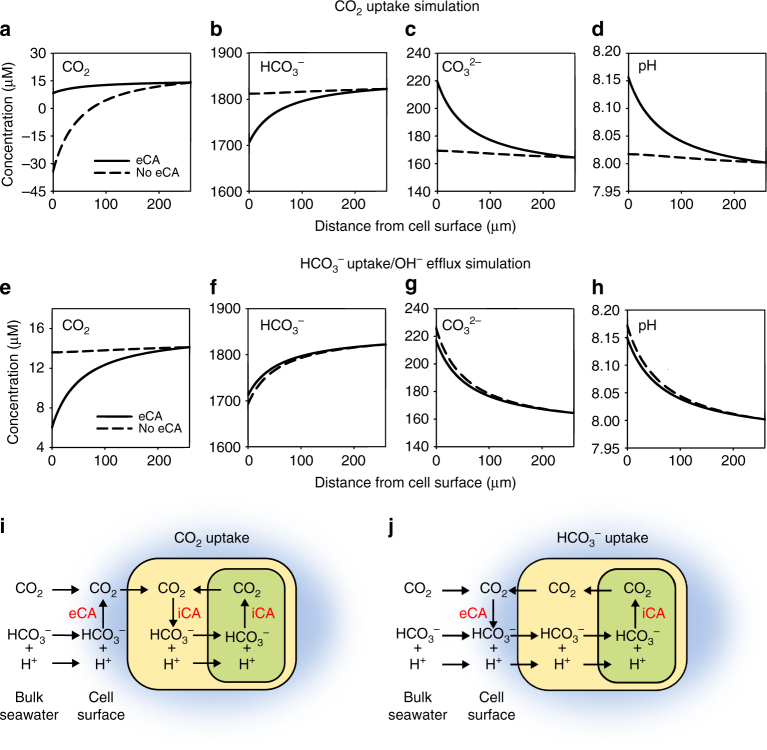
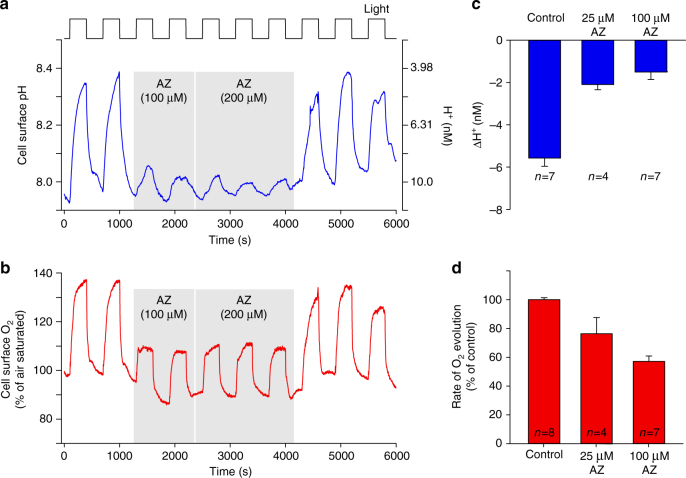
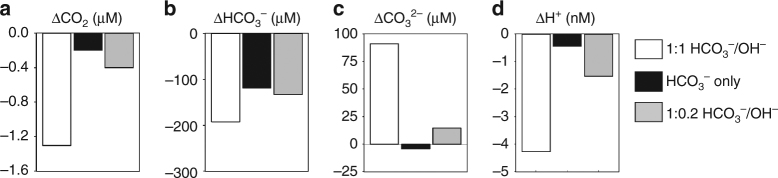
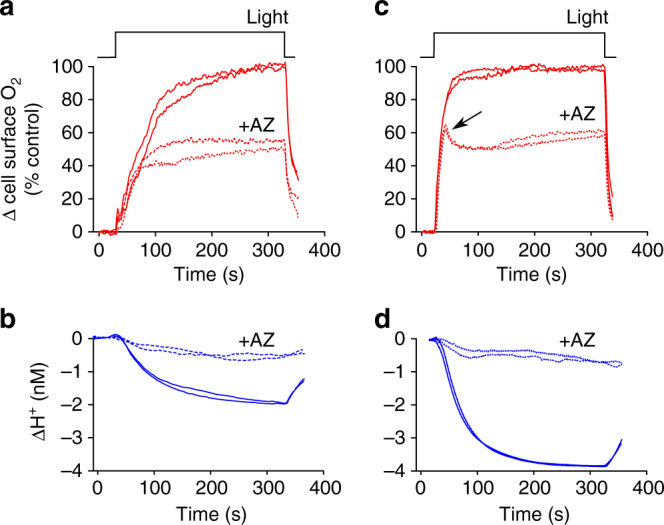
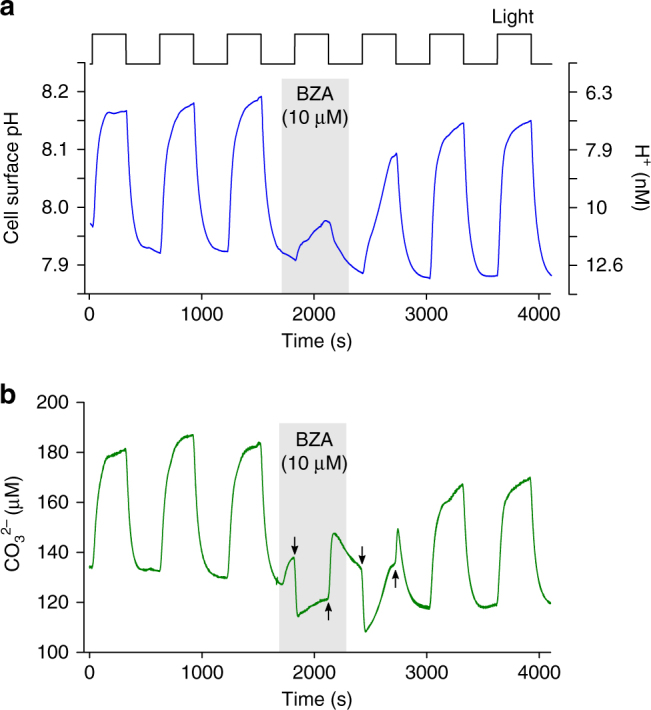
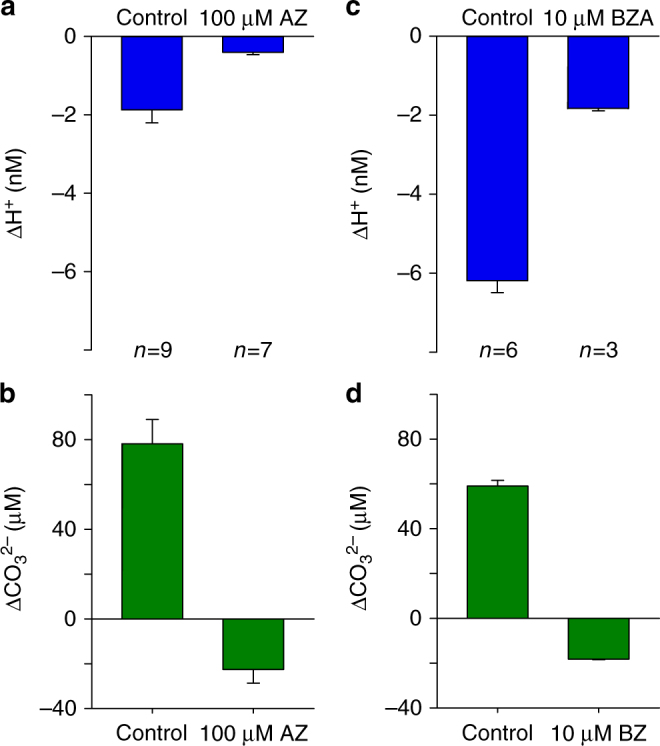
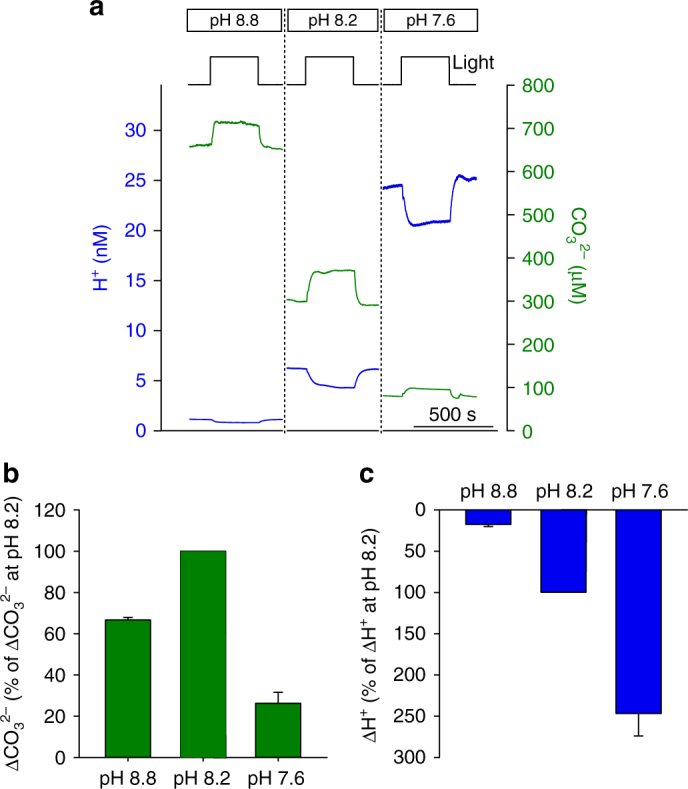
Photosynthesis by marine diatoms plays a major role in the global carbon cycle, although the precise mechanisms of dissolved inorganic carbon (DIC) uptake remain unclear. A lack of direct measurements of carbonate chemistry at the cell surface has led to uncertainty over the underlying membrane transport processes and the role of external carbonic anhydrase (eCA). Here we identify rapid and substantial photosynthesis-driven increases in pH and [CO32-] primarily due to the activity of eCA at the cell surface of the large diatom Odontella sinensis using direct simultaneous microelectrode measurements of pH and CO32- along with modelling of cell surface inorganic carbonate chemistry. Our results show that eCA acts to maintain cell surface CO2 concentrations, making a major contribution to DIC supply in O. sinensis. Carbonate chemistry at the cell surface is therefore highly dynamic and strongly dependent on cell size, morphology and the carbonate chemistry of the bulk seawater.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Nelson DM, Treguer P, Brzezinski MA, Leynaert A, Queguiner B. Production and dissolution of biogenic silica in the ocean - revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles. 1995;9:359–372. doi: 10.1029/95GB01070. - DOI
-
- Zeebe, R. & Wolf-Gladrow, D. CO2 in Seawater: Equilibrium, Kinetics, Isotopes (Elsevier, London, 2001).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
