

Deciphering the Neural Control of Sympathetic Nerve Activity: Status Report and Directions for Future Research
- PMID: 29311801
- PMCID: PMC5743742
- DOI: 10.3389/fnins.2017.00730
Deciphering the Neural Control of Sympathetic Nerve Activity: Status Report and Directions for Future Research
Abstract
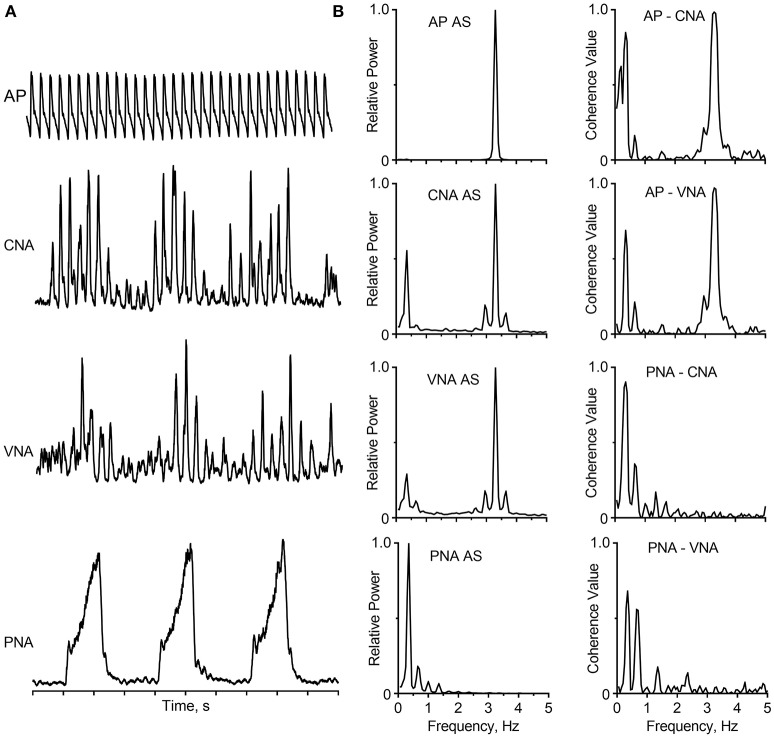
Sympathetic nerve activity (SNA) contributes appreciably to the control of physiological function, such that pathological alterations in SNA can lead to a variety of diseases. The goal of this review is to discuss the characteristics of SNA, briefly review the methodology that has been used to assess SNA and its control, and to describe the essential role of neurophysiological studies in conscious animals to provide additional insights into the regulation of SNA. Studies in both humans and animals have shown that SNA is rhythmic or organized into bursts whose frequency varies depending on experimental conditions and the species. These rhythms are generated by brainstem neurons, and conveyed to sympathetic preganglionic neurons through several pathways, including those emanating from the rostral ventrolateral medulla. Although rhythmic SNA is present in decerebrate animals (indicating that neurons in the brainstem and spinal cord are adequate to generate this activity), there is considerable evidence that a variety of supratentorial structures including the insular and prefrontal cortices, amygdala, and hypothalamic subnuclei provide inputs to the brainstem regions that regulate SNA. It is also known that the characteristics of SNA are altered during stress and particular behaviors such as the defense response and exercise. While it is a certainty that supratentorial structures contribute to changes in SNA during these behaviors, the neural underpinnings of the responses are yet to be established. Understanding how SNA is modified during affective responses and particular behaviors will require neurophysiological studies in awake, behaving animals, including those that entail recording activity from neurons that generate SNA. Recent studies have shown that responses of neurons in the central nervous system to most sensory inputs are context-specific. Future neurophysiological studies in conscious animals should also ascertain whether this general rule also applies to sensory signals that modify SNA.
Keywords: baroreceptor reflex; central command; exercise pressor response; sympathetic nerve activity; vestibulosympathetic reflex.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials