

Plant Biodiversity Drivers in Brazilian Campos Rupestres: Insights from Phylogenetic Structure
- PMID: 29312396
- PMCID: PMC5742226
- DOI: 10.3389/fpls.2017.02141
Plant Biodiversity Drivers in Brazilian Campos Rupestres: Insights from Phylogenetic Structure
Abstract
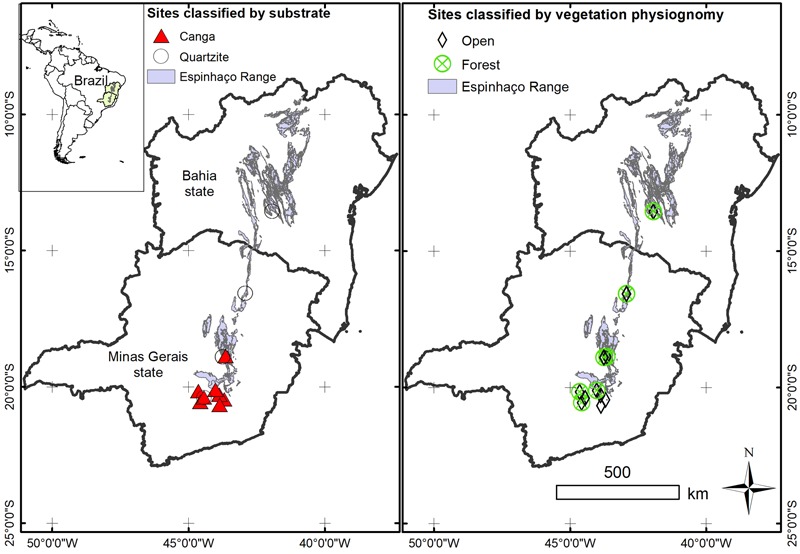
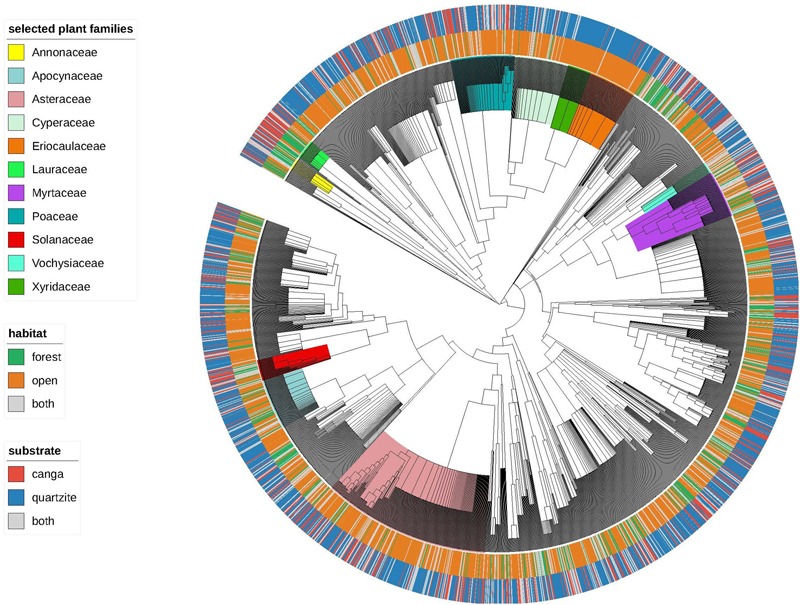
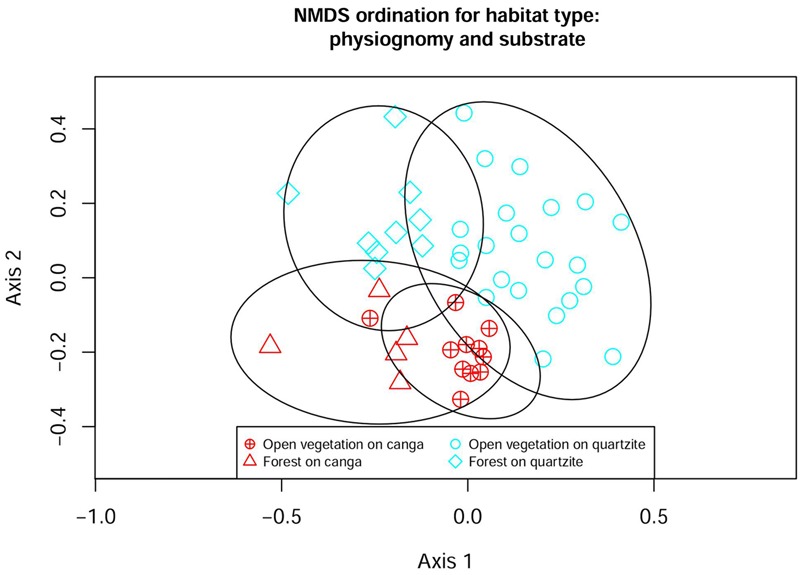
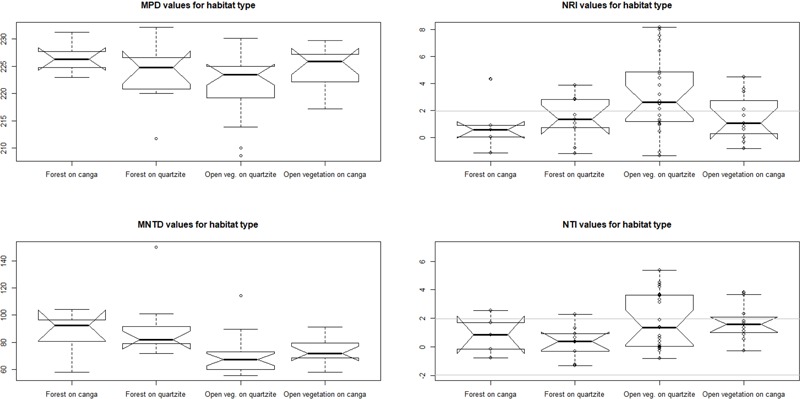
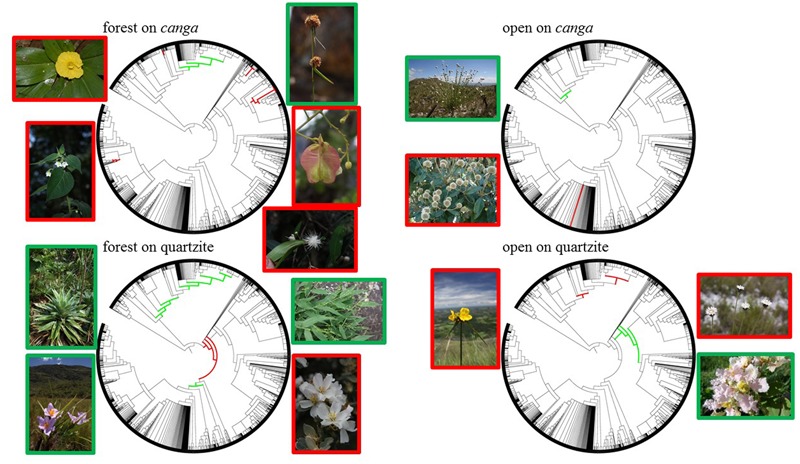
Old, climate-buffered infertile landscapes (Ocbils) have attracted increasing levels of interest in recent years because of their exceptionally diverse plant communities. Brazil's campos rupestres (rupestrian grasslands) are home to almost 15% of Brazil's native flora in less than 0.8% of Brazil's territory: an ideal study system for exploring variation in floristic diversity and phylogenetic structure in sites differing in geology and phytophysiognomy. We found significant differences in floristic diversity and phylogenetic structure across a range of study sites encompassing open vegetation and forest on quartzite (FQ) and on ironstone substrates, commonly termed canga. Substrate and physiognomy were key in structuring floristic diversity in the Espinhaço and physiognomy was more important than substrate in structuring phylogenetic diversity, with neither substrate nor its interaction with physiognomy accounting for significant variation in phylogenetic structure. Phylogenetic clustering was significant in open vegetation on both canga and quartzite, reflecting the potential role of environmental filtering in these exposed montane communities adapted to multiple environmental stressors. In forest communities, phylogenetic clustering was significant only at relatively deep nodes of the phylogeny in FQ while no significant phylogenetic clustering was detected across forest on canga (FC), which may be attributable to proximity to the megadiverse Atlantic forest biome and/or comparatively benign environmental conditions in FC with relatively deep, nutrient-rich soils and access to edaphic water reliable in comparison to those for open vegetation on canga and open or forest communities on quartzite. Clades representing relatively old lineages are significantly over-represented in campos rupestres on quartzite, consistent with the Gondwanan Heritage Hypothesis of Ocbil theory. In contrast, forested sites on canga are recognized as Yodfels. To be effective, conservation measures must take account of the distinct communities which are encompassed within the broad term campos rupestres, and the differing vulnerabilities of Ocbils and Yodfels.
Keywords: campo rupestre; canga; eastern Brazil; phylogenetic clustering; rupestrian grasslands; substrate; vegetation types.
Figures


References
-
- Alkmim F. F. (2012). “Serra do espinhaço e chapada diamantina,” in Geologia do Brasil eds Hasui Y., Carneiro C. D. R., Almeida F. F. M., Bartorelli A. (São Paulo: Editora Beca; ) 236–244.
-
- Barbosa-Silva R., Labiak P. H., Dos Santos Bragança Gil A., Goldenberg R., Michelangeli F. A., Martinelli G., et al. (2016). Over the hills and far away: new plant records for the guayana shield in Brazil. Brittonia 68 397–408. 10.1007/s12228-016-9435-3 - DOI
-
- BRAHMS (2015). University of Oxford: Botanical Research and Herbarium Management System (BRAHMS) [Online]. Available at: http://herbaria.plants.ox.ac.uk/bol/brahms [accessed November 25, 2015].
-
- Brazil Flora Group [BFG] (2015). Growing knowledge: an overview of seed plant diversity in Brazil. Rodriguésia 66 1085–1113. 10.1590/2175-7860201566411 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources