

Synthesized Heparan Sulfate Competitors Attenuate Pseudomonas aeruginosa Lung Infection
- PMID: 29315274
- PMCID: PMC5796156
- DOI: 10.3390/ijms19010207
Synthesized Heparan Sulfate Competitors Attenuate Pseudomonas aeruginosa Lung Infection
Abstract
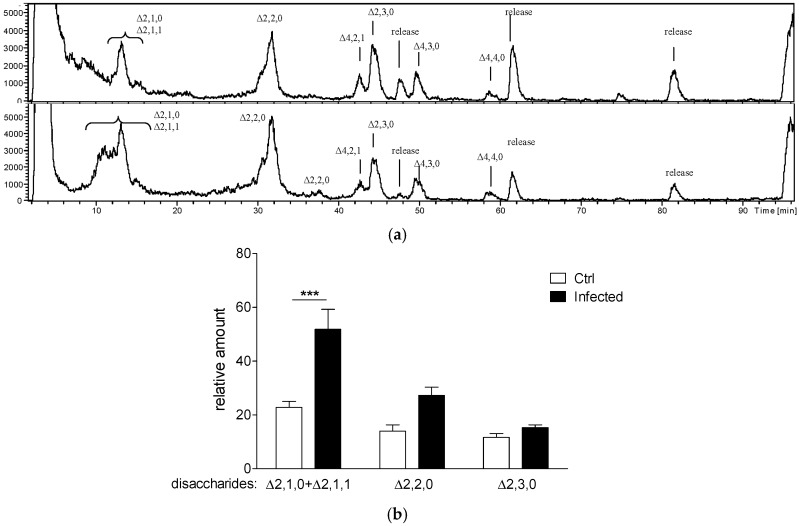
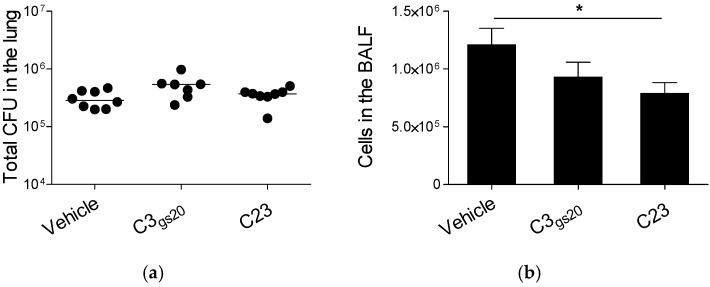
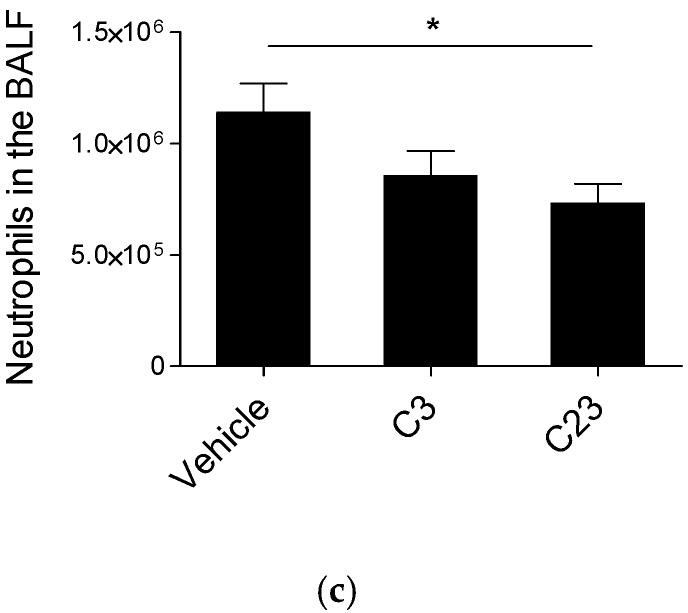
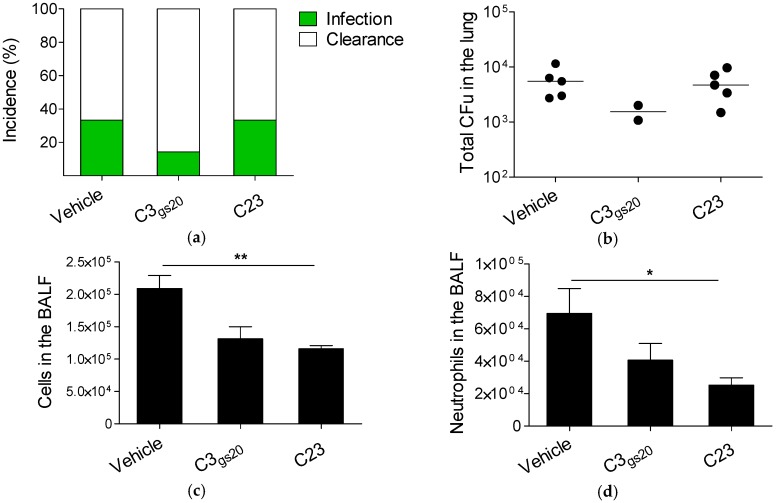
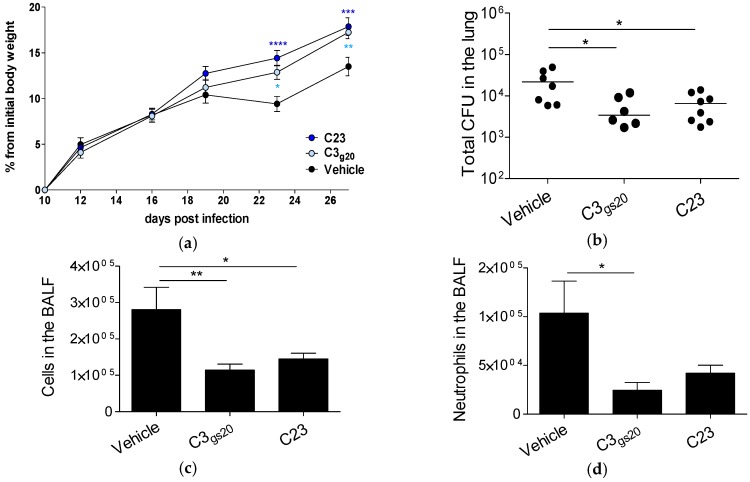
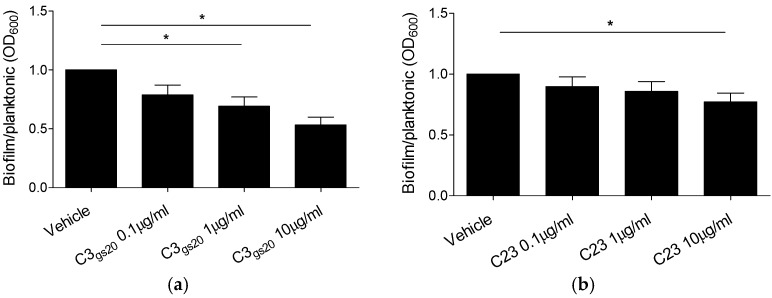
Several chronic respiratory diseases are characterized by recurrent and/or persistent infections, chronic inflammatory responses and tissue remodeling, including increased levels of glycosaminoglycans which are known structural components of the airways. Among glycosaminoglycans, heparan sulfate (HS) has been suggested to contribute to excessive inflammatory responses. Here, we aim at (i) investigating whether long-term infection by Pseudomonas aeruginosa, one of the most worrisome threat in chronic respiratory diseases, may impact HS levels, and (ii) exploring HS competitors as potential anti-inflammatory drugs during P. aeruginosa pneumonia. P. aeruginosa clinical strains and ad-hoc synthesized HS competitors were used in vitro and in murine models of lung infection. During long-term chronic P. aeruginosa colonization, infected mice showed higher heparin/HS levels, evaluated by high performance liquid chromatography-mass spectrometry after selective enzymatic digestion, compared to uninfected mice. Among HS competitors, an N-acetyl heparin and a glycol-split heparin dampened leukocyte recruitment and cytokine/chemokine production induced by acute and chronic P. aeruginosa pneumonia in mice. Furthermore, treatment with HS competitors reduced bacterial burden during chronic murine lung infection. In vitro, P. aeruginosa biofilm formation decreased upon treatment with HS competitors. Overall, these findings support further evaluation of HS competitors as a novel therapy to counteract inflammation and infection during P. aeruginosa pneumonia.
Keywords: Pseudomonas aeruginosa infections; anti-inflammatory drugs; chronic respiratory diseases; glycosaminoglycans; mouse models.
Conflict of interest statement
The authors declare no conflict of interest. The founding sponsor had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Figures
References
-
- Proudfoot A.E., Handel T.M., Johnson Z., Lau E.K., LiWang P., Clark-Lewis I., Borlat F., Wells T.N., Kosco-Vilbois M.H. Glycosaminoglycan binding and oligomerization are essential for the in vivo activity of certain chemokines. Proc. Natl. Acad. Sci. USA. 2003;100:1885–1890. doi: 10.1073/pnas.0334864100. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
