

The Role of Hippocampal Replay in Memory and Planning
- PMID: 29316421
- PMCID: PMC5847173
- DOI: 10.1016/j.cub.2017.10.073
The Role of Hippocampal Replay in Memory and Planning
Abstract
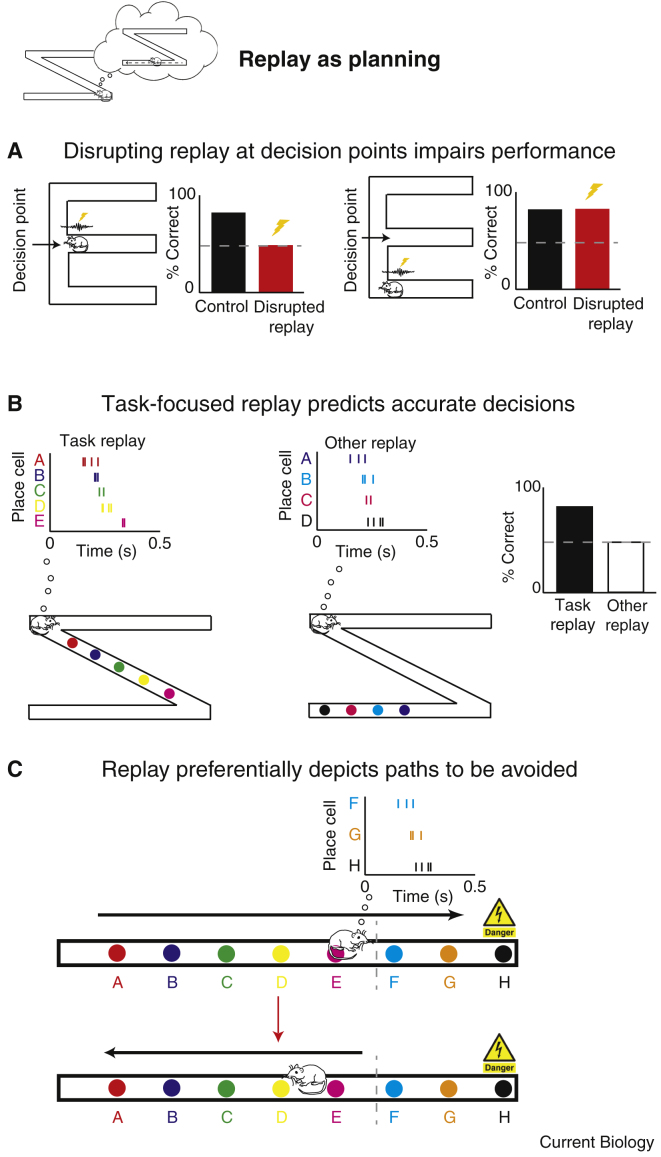
The mammalian hippocampus is important for normal memory function, particularly memory for places and events. Place cells, neurons within the hippocampus that have spatial receptive fields, represent information about an animal's position. During periods of rest, but also during active task engagement, place cells spontaneously recapitulate past trajectories. Such 'replay' has been proposed as a mechanism necessary for a range of neurobiological functions, including systems memory consolidation, recall and spatial working memory, navigational planning, and reinforcement learning. Focusing mainly, but not exclusively, on work conducted in rodents, we describe the methodologies used to analyse replay and review evidence for its putative roles. We identify outstanding questions as well as apparent inconsistencies in existing data, making suggestions as to how these might be resolved. In particular, we find support for the involvement of replay in disparate processes, including the maintenance of hippocampal memories and decision making. We propose that the function of replay changes dynamically according to task demands placed on an organism and its current level of arousal.
Copyright © 2017 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- O'Keefe J., Nadel L. Clarendon; Oxford: 1978. The Hippocampus as a Cognitive Map.
-
- Teng E., Squire L.R. Memory for places learned long ago is intact after hippocampal damage. Nature. 1999;400:675–677. - PubMed
-
- Cipolotti L., Shallice T., Chan D., Fox N., Scahill R., Harrison G., Stevens J., Rudge P. Long-term retrograde amnesia...the crucial role of the hippocampus. Neuropsychologia. 2001;39:151–172. - PubMed
-
- Rosenbaum R.S., Priselac S., Kohler S., Black S.E., Gao F., Nadel L., Moscovitch M. Remote spatial memory in an amnesic person with extensive bilateral hippocampal lesions. Nat. Neurosci. 2000;3:1044–1048. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
