

A mutation in Ccdc39 causes neonatal hydrocephalus with abnormal motile cilia development in mice
- PMID: 29317443
- PMCID: PMC5825874
- DOI: 10.1242/dev.154500
A mutation in Ccdc39 causes neonatal hydrocephalus with abnormal motile cilia development in mice
Abstract
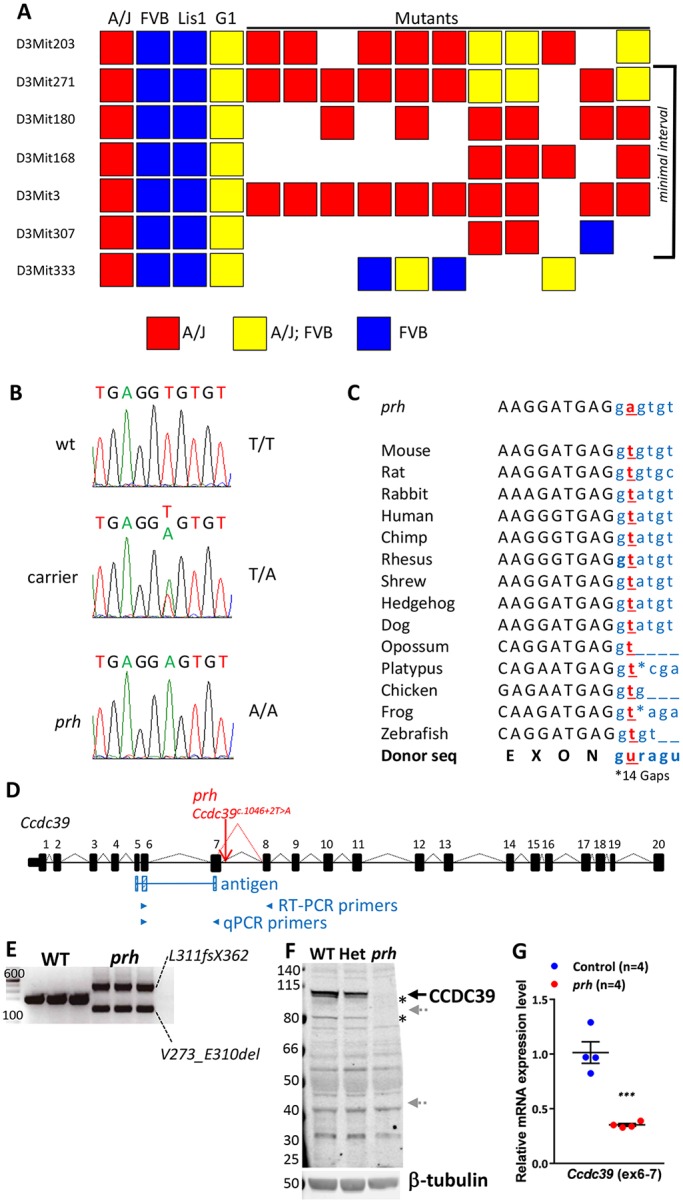
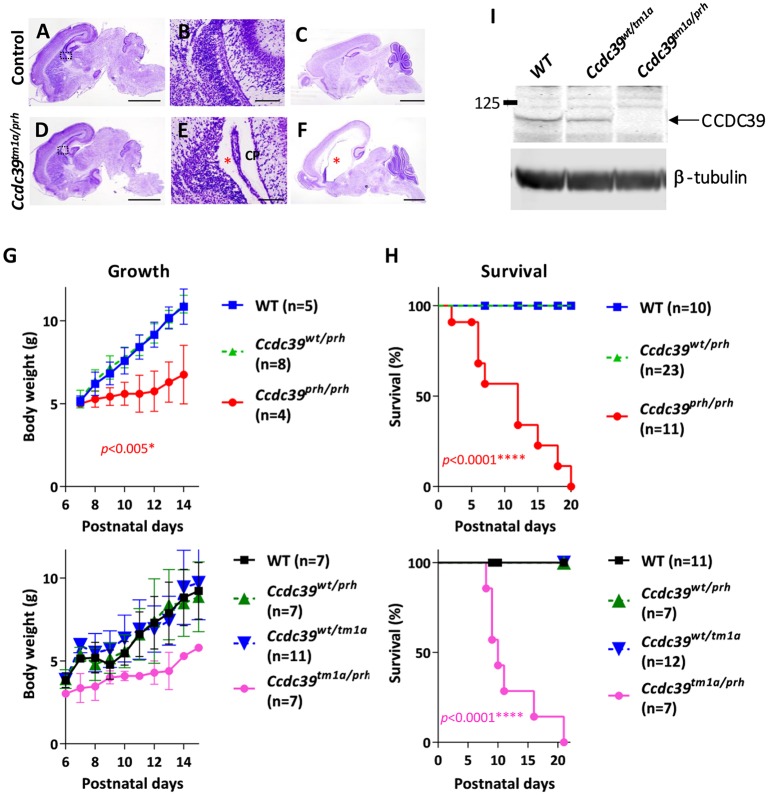
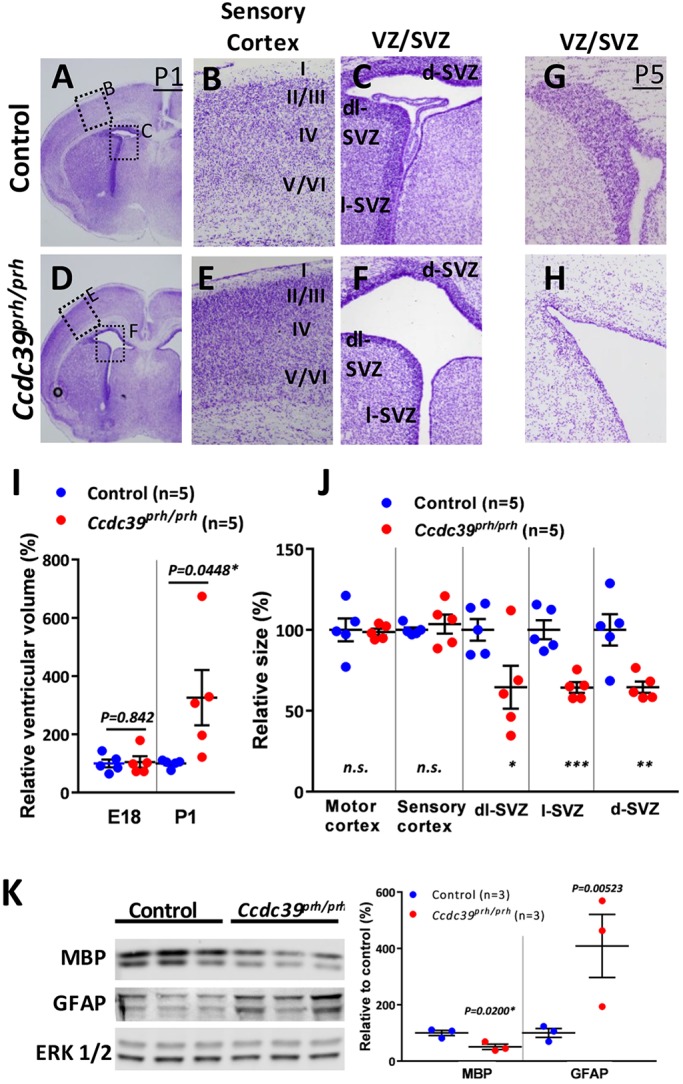
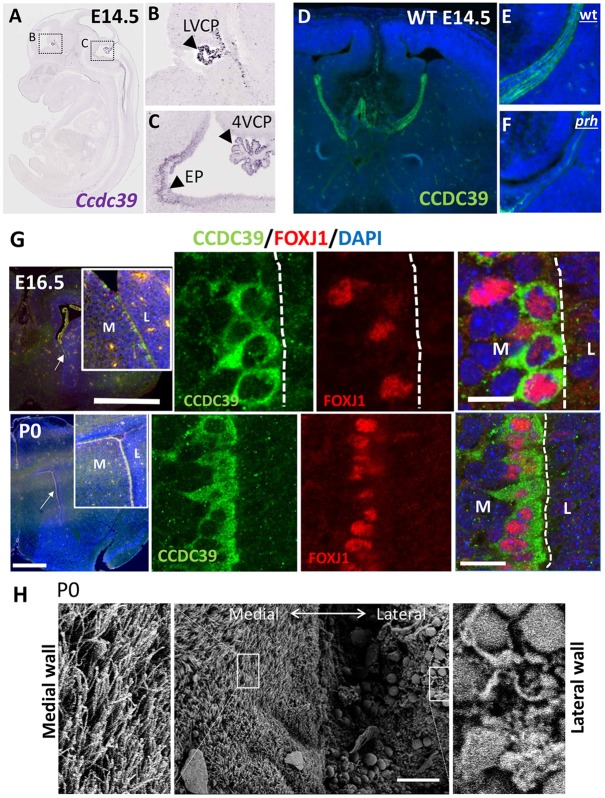
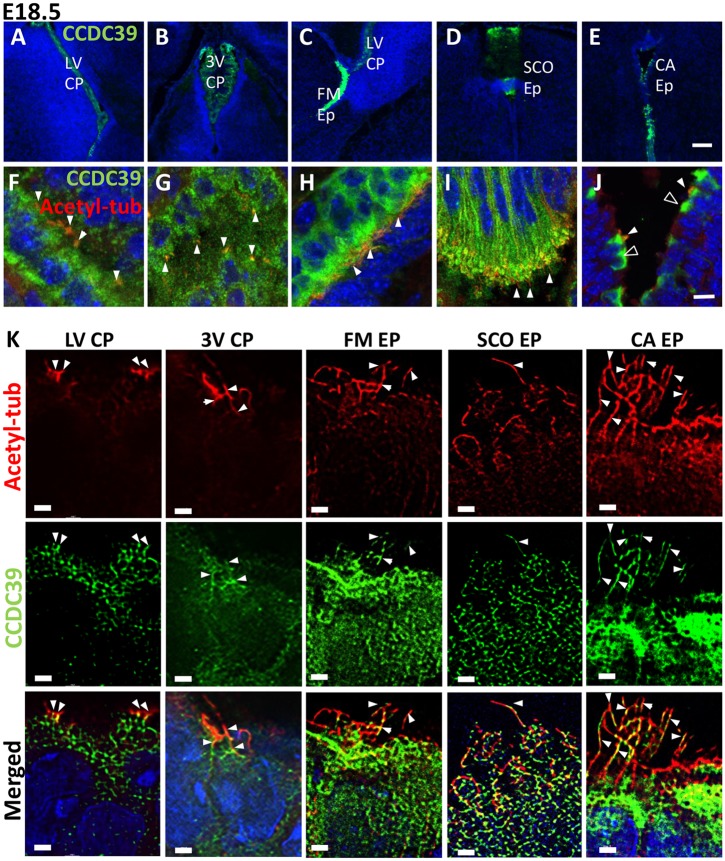
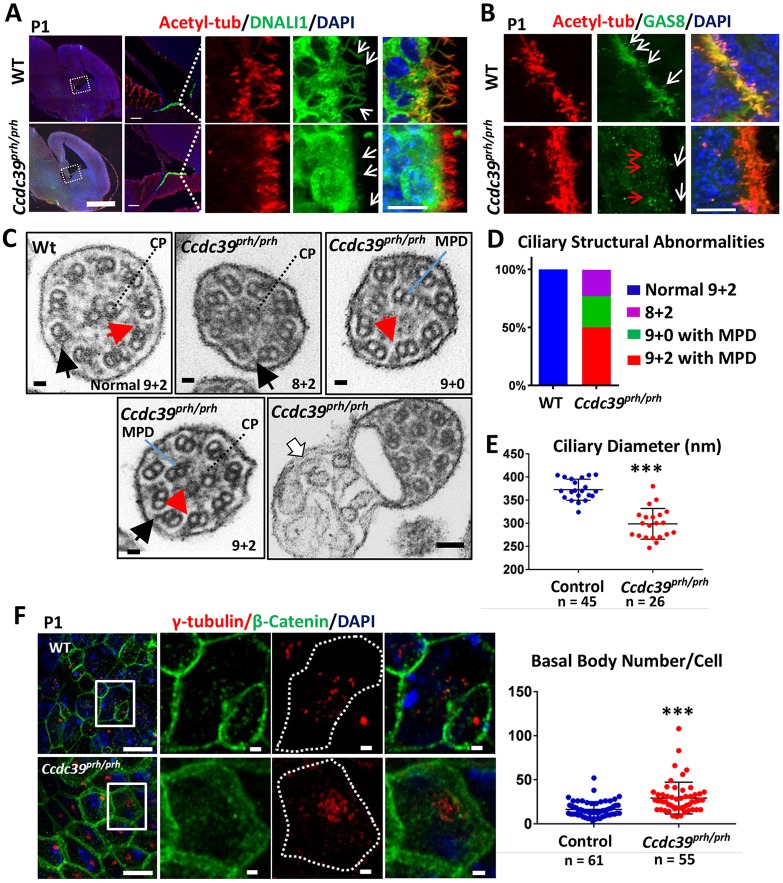
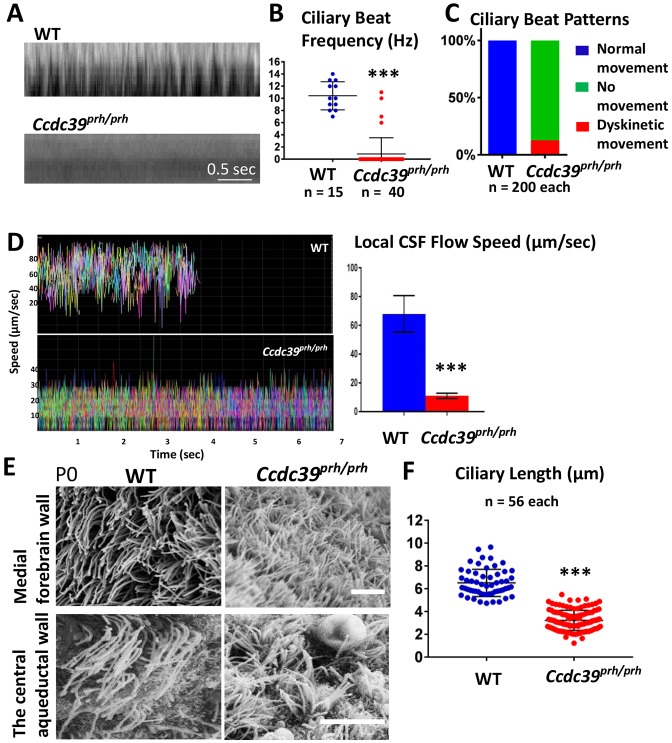
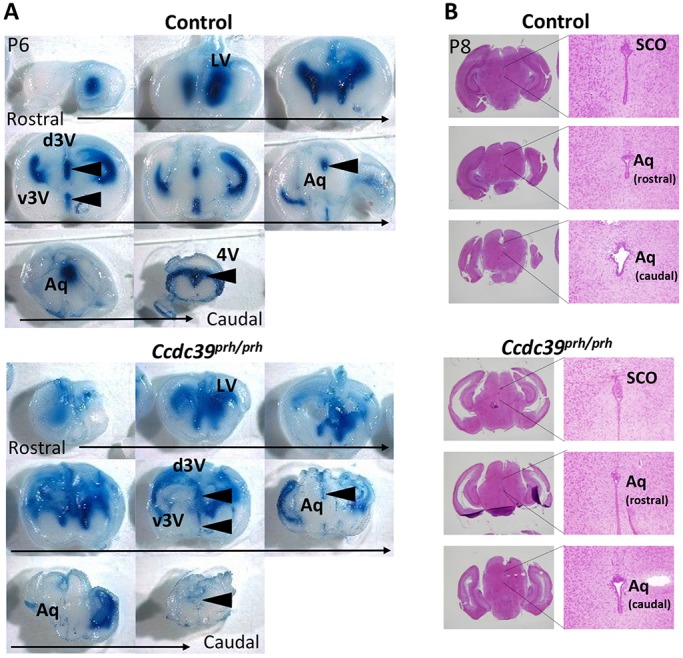
Pediatric hydrocephalus is characterized by an abnormal accumulation of cerebrospinal fluid (CSF) and is one of the most common congenital brain abnormalities. However, little is known about the molecular and cellular mechanisms regulating CSF flow in the developing brain. Through whole-genome sequencing analysis, we report that a homozygous splice site mutation in coiled-coil domain containing 39 (Ccdc39) is responsible for early postnatal hydrocephalus in the progressive hydrocephalus (prh) mouse mutant. Ccdc39 is selectively expressed in embryonic choroid plexus and ependymal cells on the medial wall of the forebrain ventricle, and the protein is localized to the axoneme of motile cilia. The Ccdc39prh/prh ependymal cells develop shorter cilia with disorganized microtubules lacking the axonemal inner arm dynein. Using high-speed video microscopy, we show that an orchestrated ependymal ciliary beating pattern controls unidirectional CSF flow on the ventricular surface, which generates bulk CSF flow in the developing brain. Collectively, our data provide the first evidence for involvement of Ccdc39 in hydrocephalus and suggest that the proper development of medial wall ependymal cilia is crucial for normal mouse brain development.
Keywords: Brain development; Cerebrospinal fluid; Cilia; Ependymal cells; Hydrocephalus.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Antony D., Becker-Heck A., Zariwala M. A., Schmidts M., Onoufriadis A., Forouhan M., Wilson R., Taylor-Cox T., Dewar A., Jackson C. et al. (2013). Mutations in CCDC39 and CCDC40 are the major cause of primary ciliary dyskinesia with axonemal disorganization and absent inner dynein arms. Hum. Mutat. 34, 462-472. 10.1002/humu.22261 - DOI - PMC - PubMed
-
- Banizs B., Pike M. M., Millican C. L., Ferguson W. B., Komlosi P., Sheetz J., Bell P. D., Schwiebert E. M. and Yoder B. K. (2005). Dysfunctional cilia lead to altered ependyma and choroid plexus function, and result in the formation of hydrocephalus. Development 132, 5329-5339. 10.1242/dev.02153 - DOI - PubMed
-
- Banizs B., Komlosi P., Bevensee M. O., Schwiebert E. M., Bell P. D. and Yoder B. K. (2007). Altered pH(i) regulation and Na(+)/HCO3(−) transporter activity in choroid plexus of cilia-defective Tg737(orpk) mutant mouse. Am. J. Physiol. Cell Physiol. 292, C1409-C1416. 10.1152/ajpcell.00408.2006 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
