

The relationship between form and function throughout the history of excitation-contraction coupling
- PMID: 29317466
- PMCID: PMC5806676
- DOI: 10.1085/jgp.201711889
The relationship between form and function throughout the history of excitation-contraction coupling
Erratum in
-
Correction: The relationship between form and function throughout the history of excitation-contraction coupling.J Gen Physiol. 2018 Feb 5;150(2):369. doi: 10.1085/jgp.20171188901162018c. Epub 2018 Jan 22. J Gen Physiol. 2018. PMID: 29358187 Free PMC article. No abstract available.
Abstract
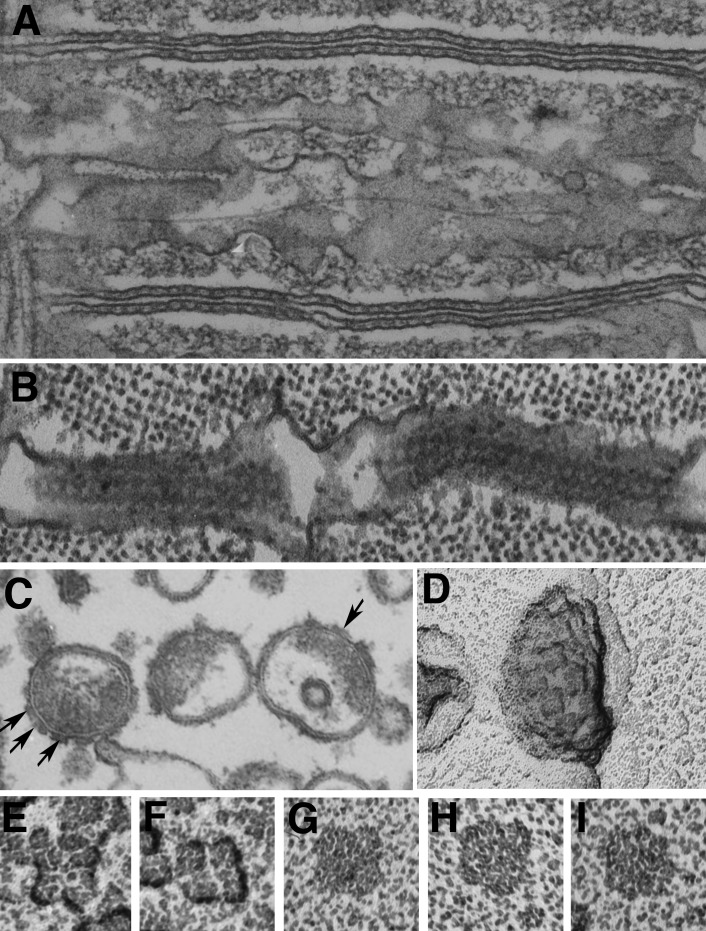
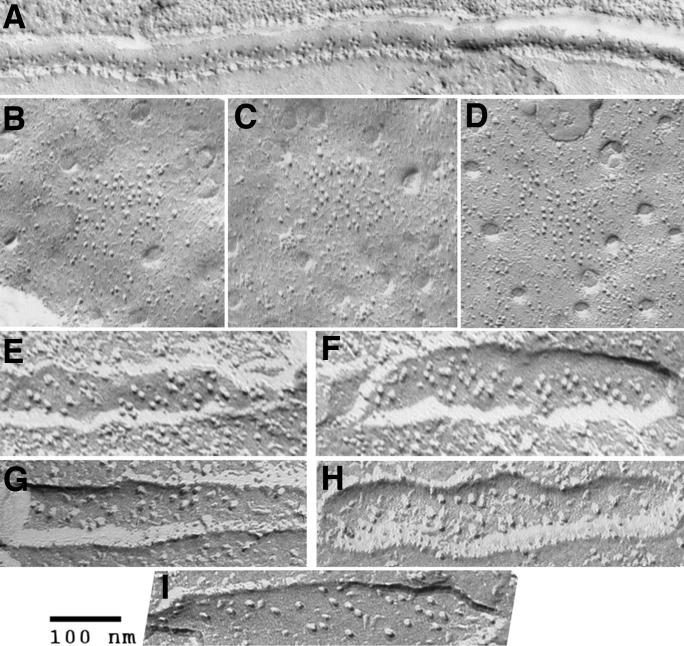
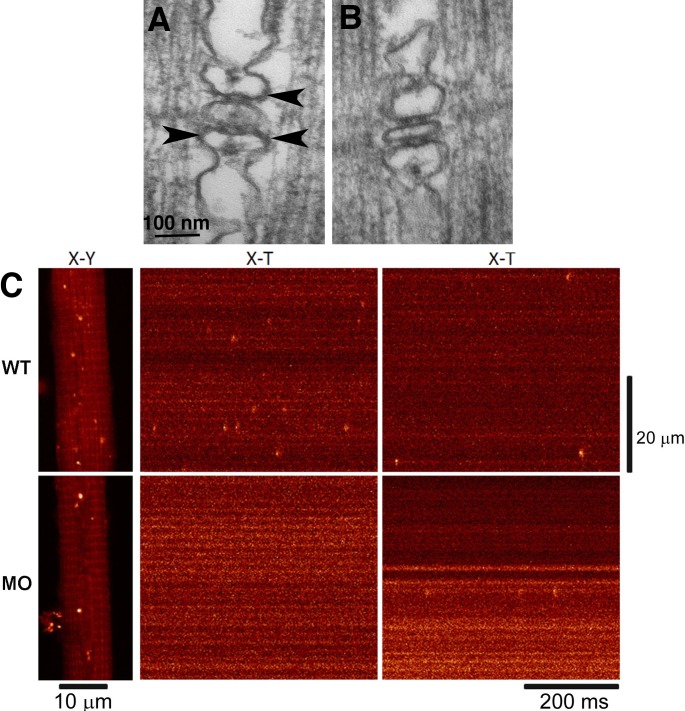
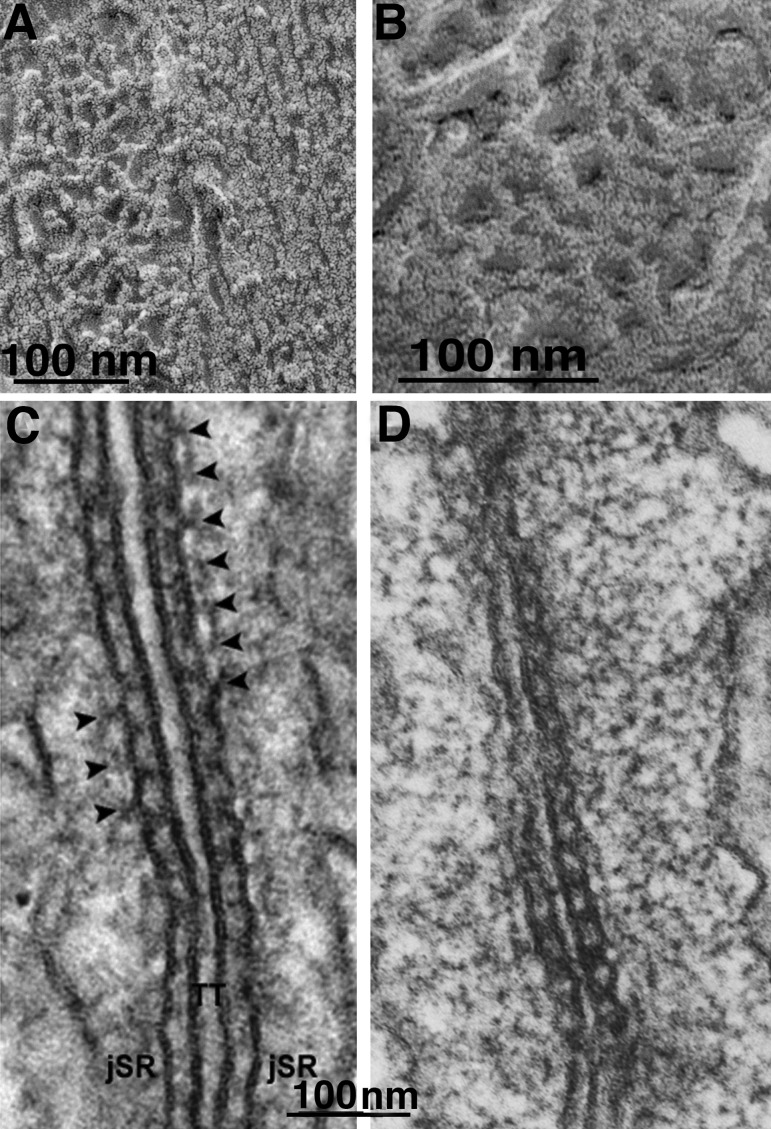
The concept of excitation-contraction coupling is almost as old as Journal of General Physiology It was understood as early as the 1940s that a series of stereotyped events is responsible for the rapid contraction response of muscle fibers to an initial electrical event at the surface. These early developments, now lost in what seems to be the far past for most young investigators, have provided an endless source of experimental approaches. In this Milestone in Physiology, I describe in detail the experiments and concepts that introduced and established the field of excitation-contraction coupling in skeletal muscle. More recent advances are presented in an abbreviated form, as readers are likely to be familiar with recent work in the field.
© 2018 Franzini-Armstrong.
Figures
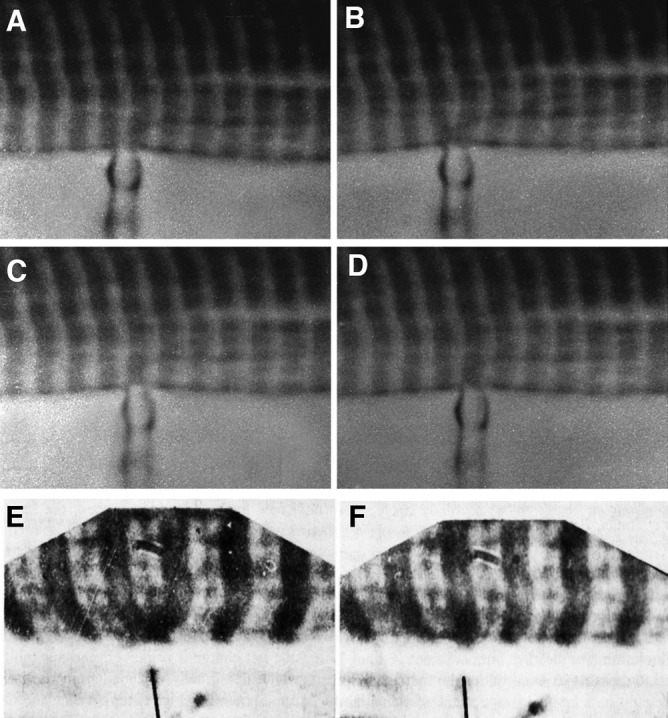
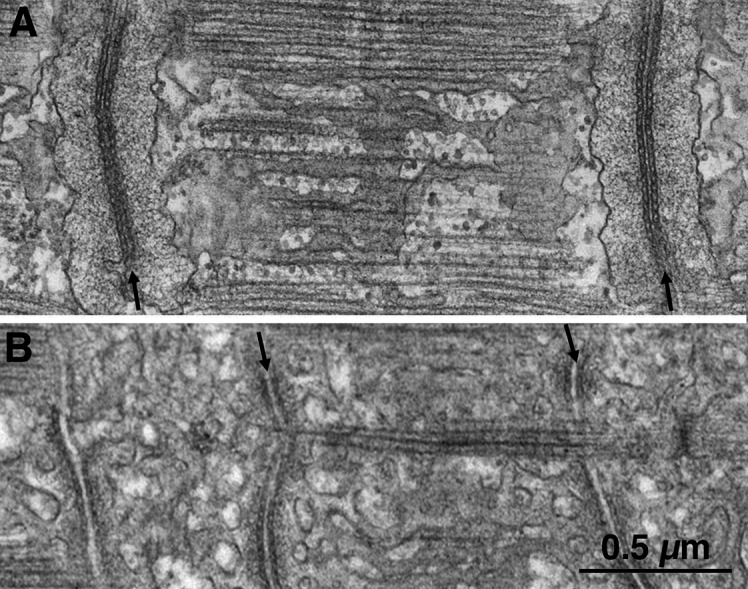

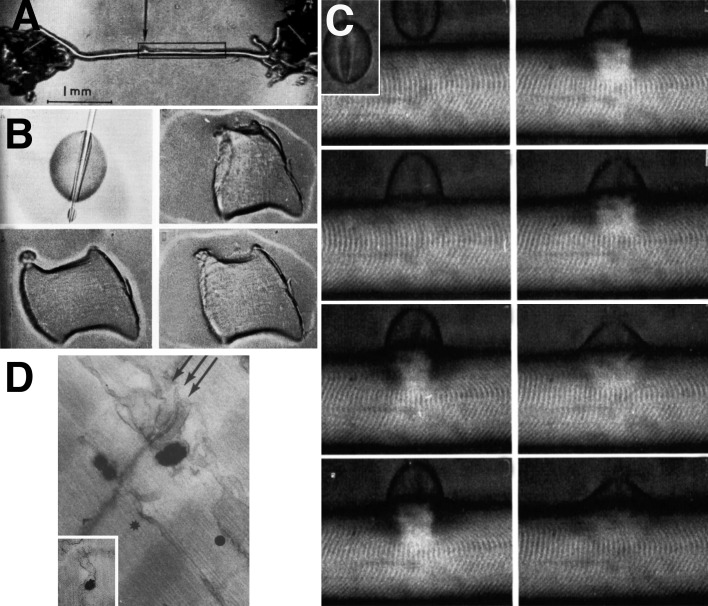
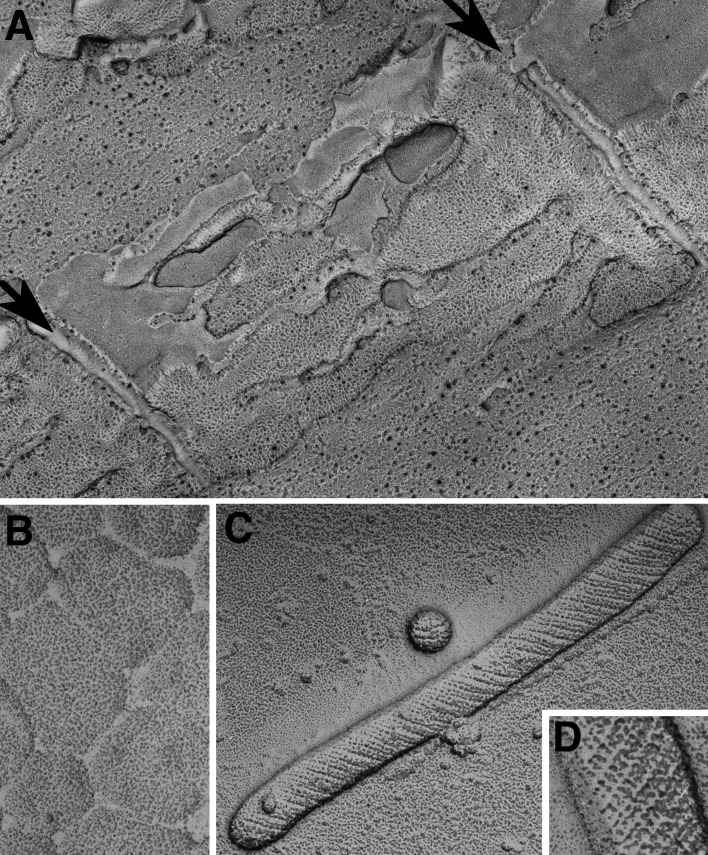

References
-
- Almers W. 1980. Potassium concentration changes in the transverse tubules of vertebrate skeletal muscle. Fed. Proc. 39:1527–1532. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
