

Maternal Adaptive Immune Cells in Decidua Parietalis Display a More Activated and Coinhibitory Phenotype Compared to Decidua Basalis
- PMID: 29317870
- PMCID: PMC5727765
- DOI: 10.1155/2017/8010961
Maternal Adaptive Immune Cells in Decidua Parietalis Display a More Activated and Coinhibitory Phenotype Compared to Decidua Basalis
Abstract
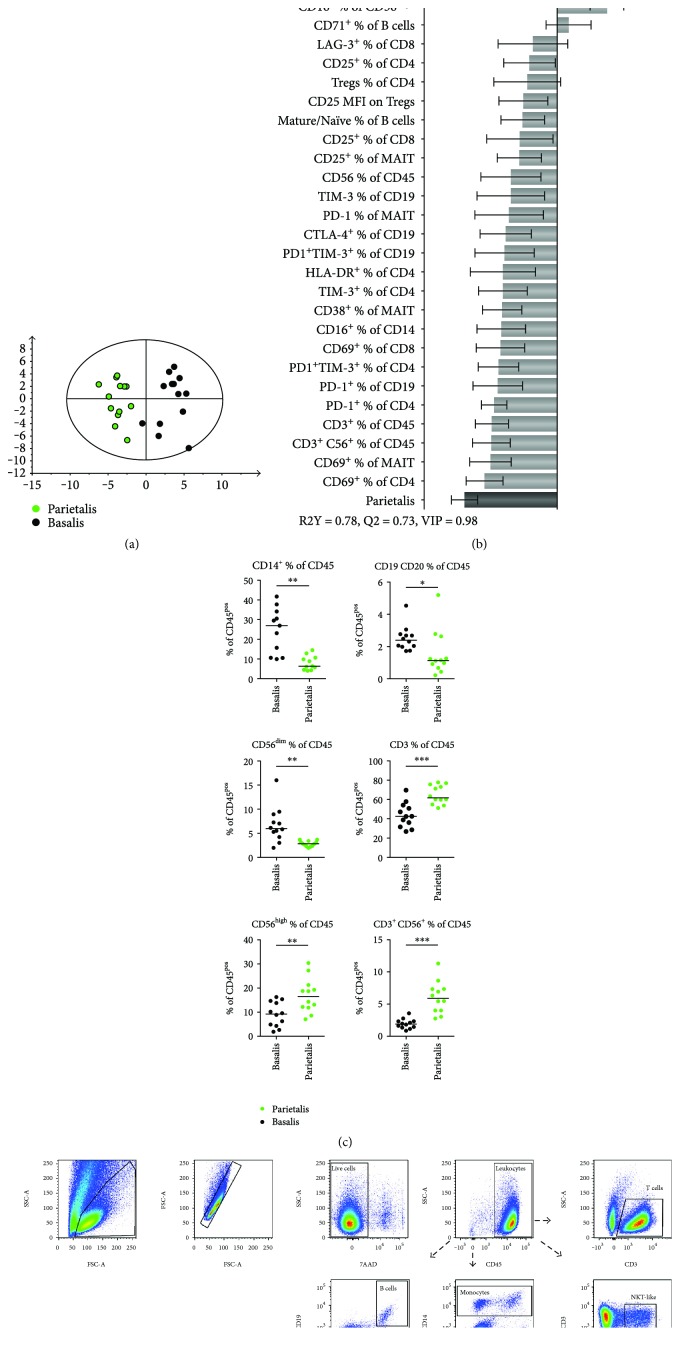
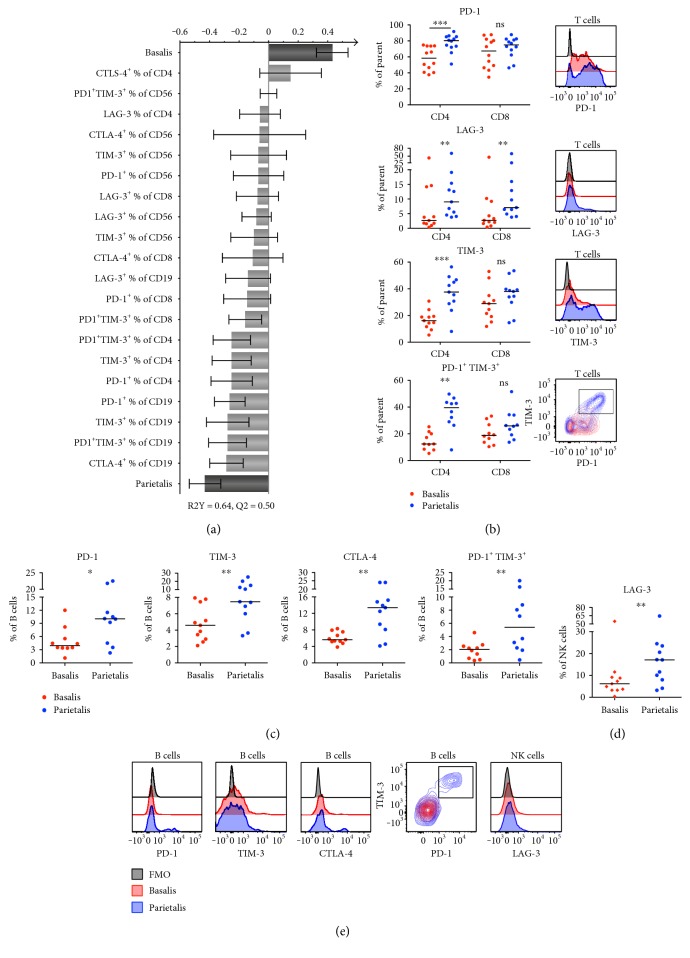
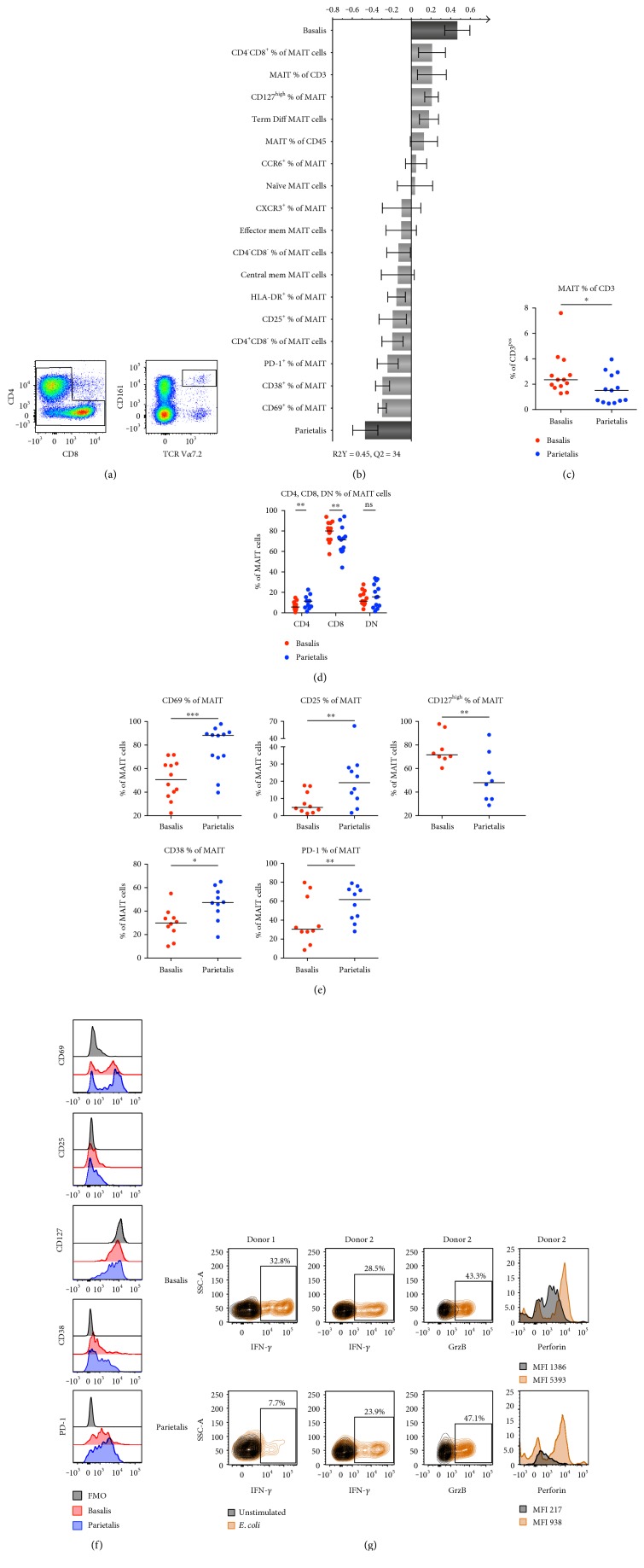
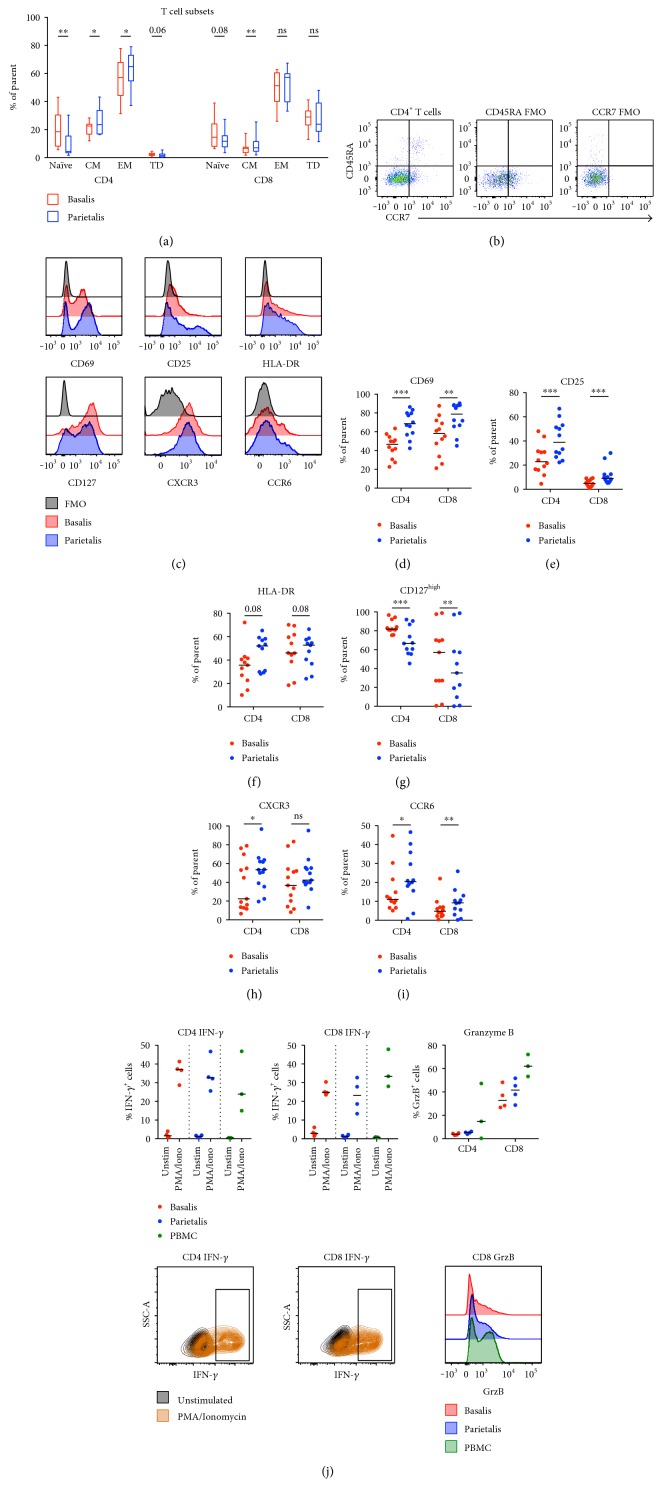
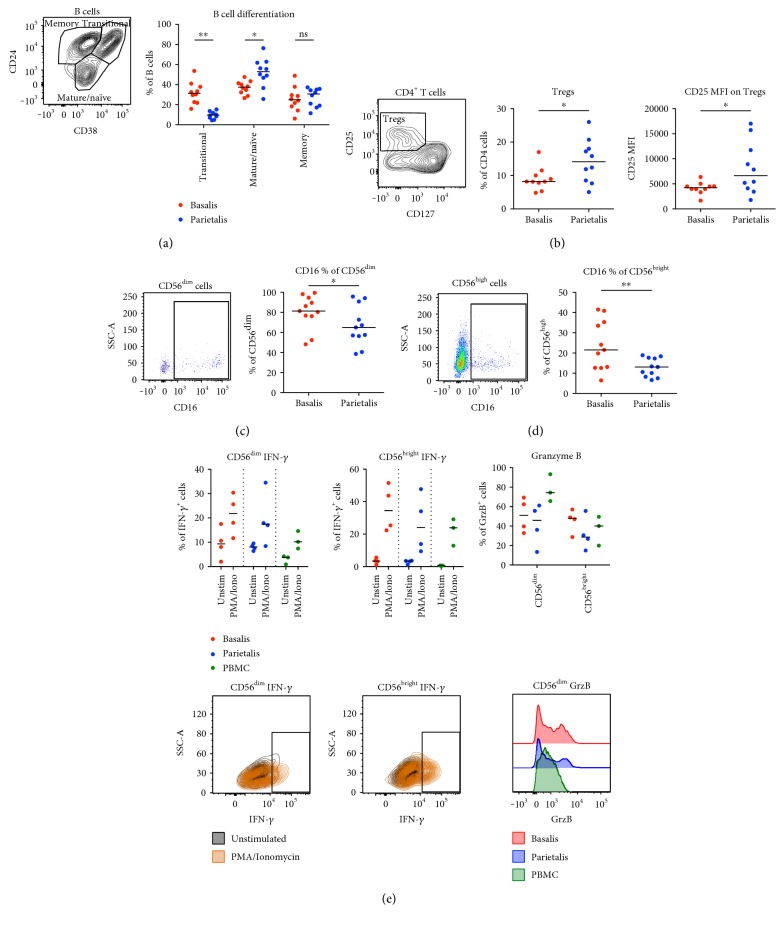
The maternal part of the placenta, the decidua, consists of maternal immune cells, decidual stromal cells, and extravillous fetal trophoblasts. In a successful pregnancy, these cell compartments interact to provide an intricate balance between fetal tolerance and antimicrobial defense. These processes are still poorly characterized in the two anatomically different decidual tissues, basalis and parietalis. We examined immune cells from decidua basalis and parietalis from term placentas (n = 15) with flow cytometry. By using multivariate discriminant analysis, we found a clear separation between the two decidual compartments based on the 81 investigated parameters. Decidua parietalis lymphocytes displayed a more activated phenotype with a higher expression of coinhibitory markers than those isolated from basalis and contained higher frequencies of T regulatory cells. Decidua basalis contained higher proportions of monocytes, B cells, and mucosal-associated invariant T (MAIT) cells. The basalis B cells were more immature, and parietalis MAIT cells showed a more activated phenotype. Conventional T cells, NK cells, and MAIT cells from both compartments potently responded with the production of interferon-γ and/or cytotoxic molecules in response to stimulation. To conclude, leukocytes in decidua basalis and parietalis displayed remarkable phenotypic disparities, indicating that the corresponding stromal microenvironments provide different immunoregulatory signals.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
