

Transcription control engineering and applications in synthetic biology
- PMID: 29318198
- PMCID: PMC5655343
- DOI: 10.1016/j.synbio.2017.09.003
Transcription control engineering and applications in synthetic biology
Abstract
In synthetic biology, researchers assemble biological components in new ways to produce systems with practical applications. One of these practical applications is control of the flow of genetic information (from nucleic acid to protein), a.k.a. gene regulation. Regulation is critical for optimizing protein (and therefore activity) levels and the subsequent levels of metabolites and other cellular properties. The central dogma of molecular biology posits that information flow commences with transcription, and accordingly, regulatory tools targeting transcription have received the most attention in synthetic biology. In this mini-review, we highlight many past successes and summarize the lessons learned in developing tools for controlling transcription. In particular, we focus on engineering studies where promoters and transcription terminators (cis-factors) were directly engineered and/or isolated from DNA libraries. We also review several well-characterized transcription regulators (trans-factors), giving examples of how cis- and trans-acting factors have been combined to create digital and analogue switches for regulating transcription in response to various signals. Last, we provide examples of how engineered transcription control systems have been used in metabolic engineering and more complicated genetic circuits. While most of our mini-review focuses on the well-characterized bacterium Escherichia coli, we also provide several examples of the use of transcription control engineering in non-model organisms. Similar approaches have been applied outside the bacterial kingdom indicating that the lessons learned from bacterial studies may be generalized for other organisms.
Keywords: Bacteria; Metabolic engineering; Promoter; Synthetic biology; Termination; Transcription.
Figures
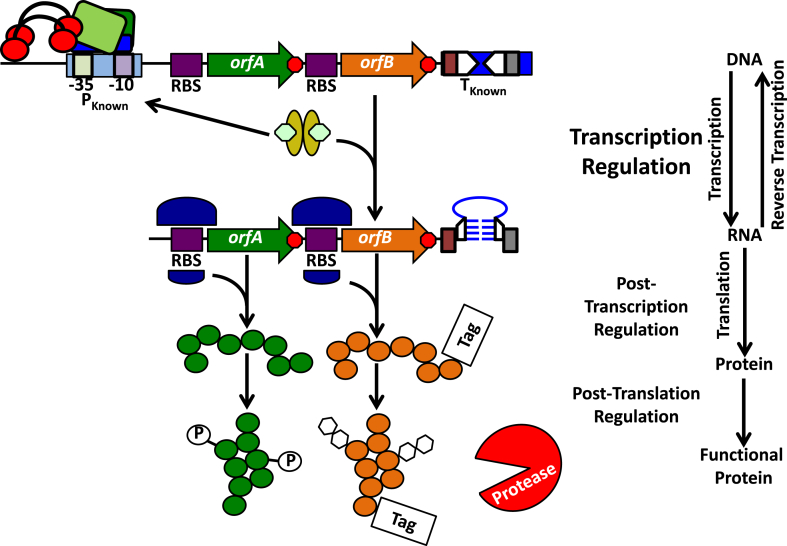
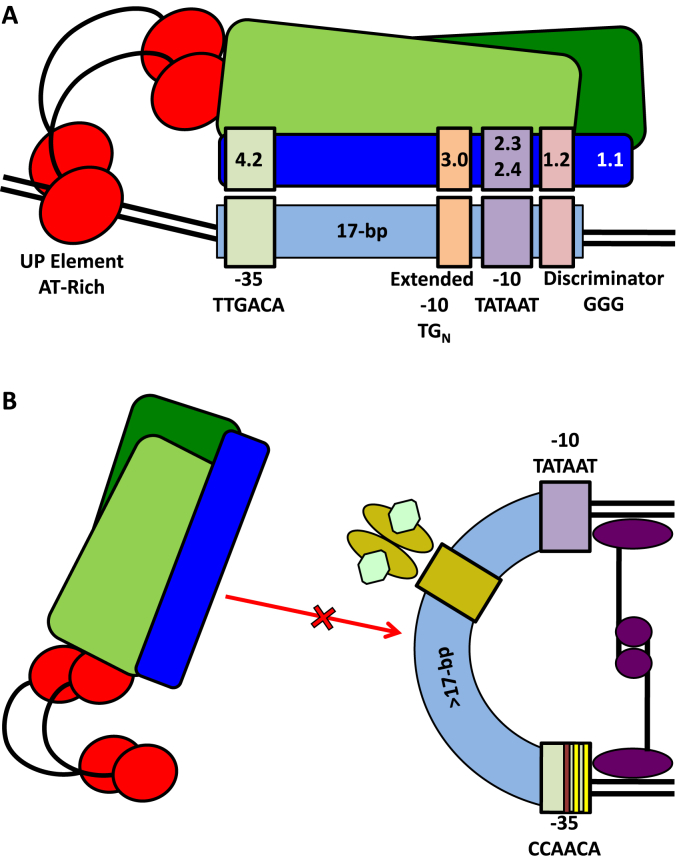
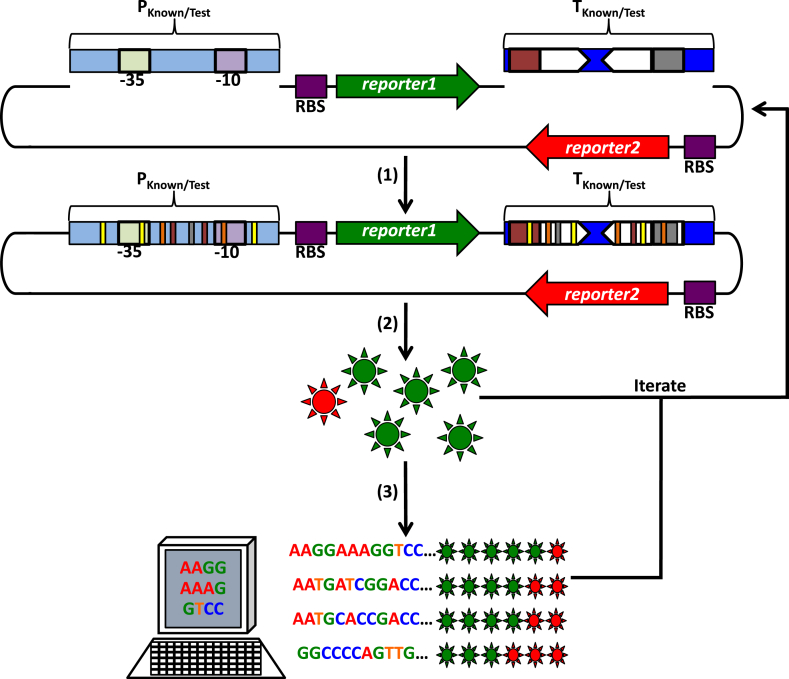
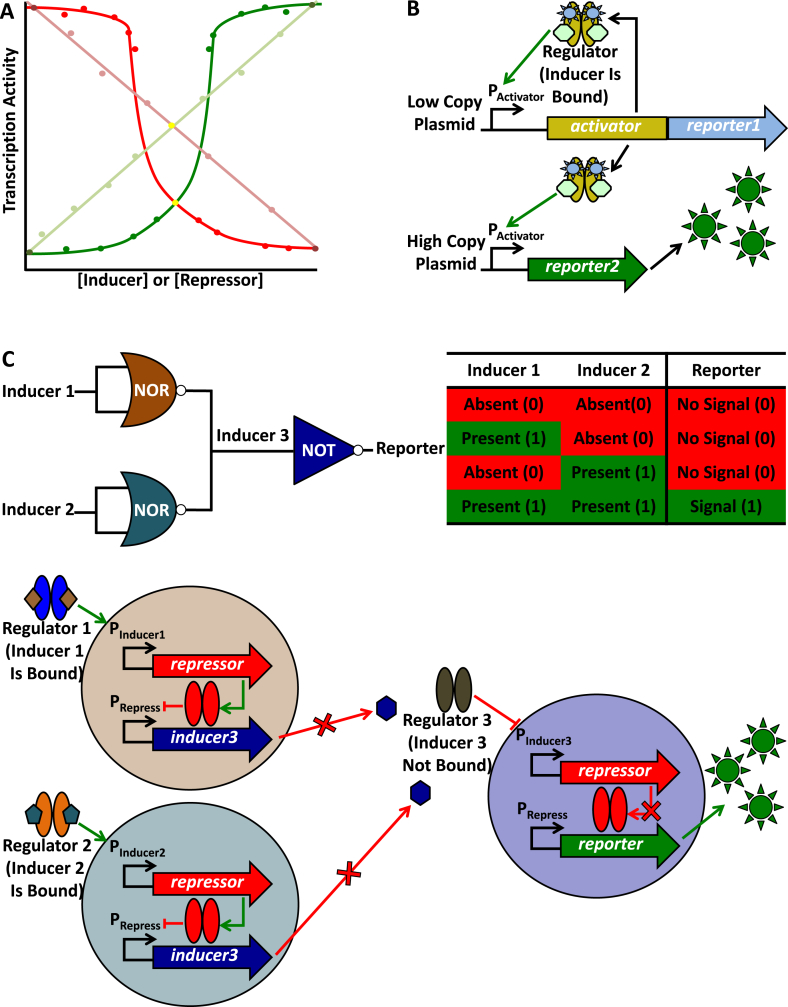
References
-
- Crick F. Central dogma of molecular biology. Nature. 1970;227:561–563. - PubMed
-
- Cech T.R. Structural biology. The ribosome is a ribozyme. Science. 2000;289:878–879. - PubMed
-
- Ban N., Nissen P., Hansen J., Moore P.B., Steitz T.A. The complete atomic structure of the large ribosomal subunit at 2.4 A resolution. Science. 2000;289:905–920. - PubMed
-
- Nissen P., Hansen J., Ban N., Moore P.B., Steitz T.A. The structural basis of ribosome activity in peptide bond synthesis. Science. 2000;289:920–930. - PubMed
-
- Muth G.W., Ortoleva-Donnelly L., Strobel S.A. A single adenosine with a neutral pKa in the ribosomal peptidyl transferase center. Science. 2000;289:947–950. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
