

Pharmacological activation of REV-ERBs is lethal in cancer and oncogene-induced senescence
- PMID: 29320480
- PMCID: PMC5924733
- DOI: 10.1038/nature25170
Pharmacological activation of REV-ERBs is lethal in cancer and oncogene-induced senescence
Abstract
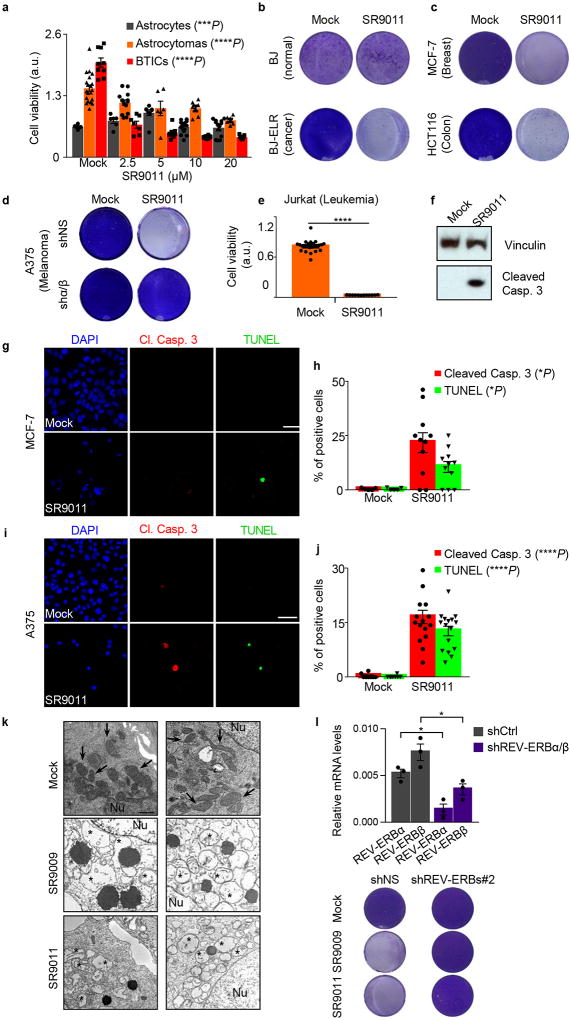
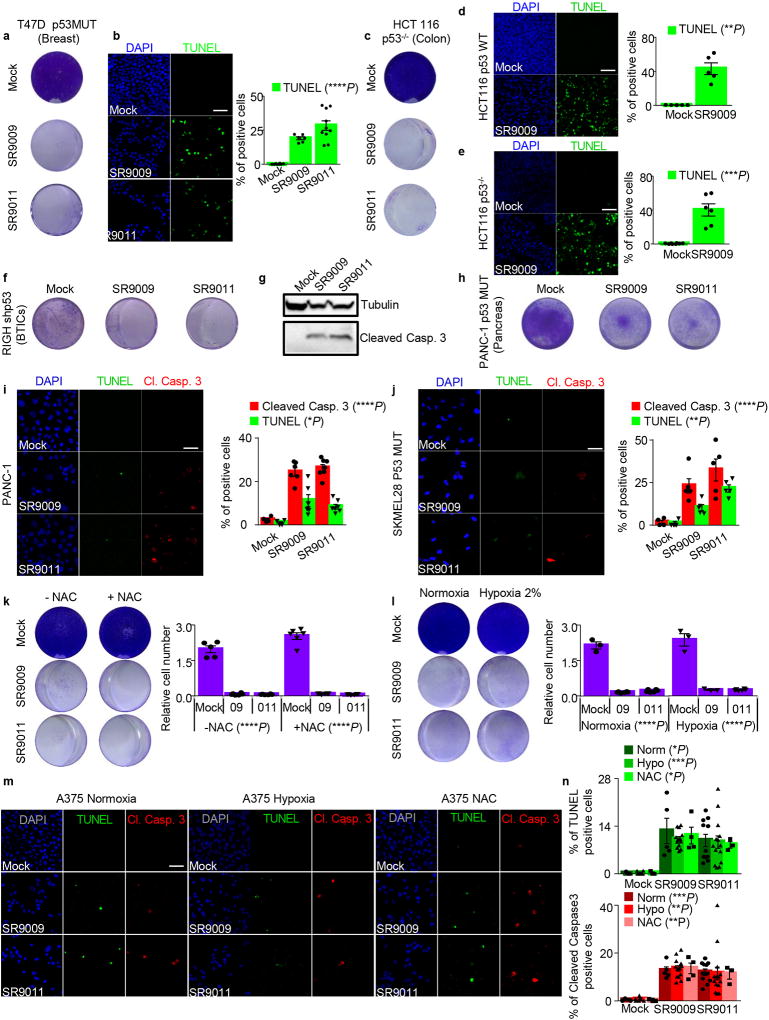
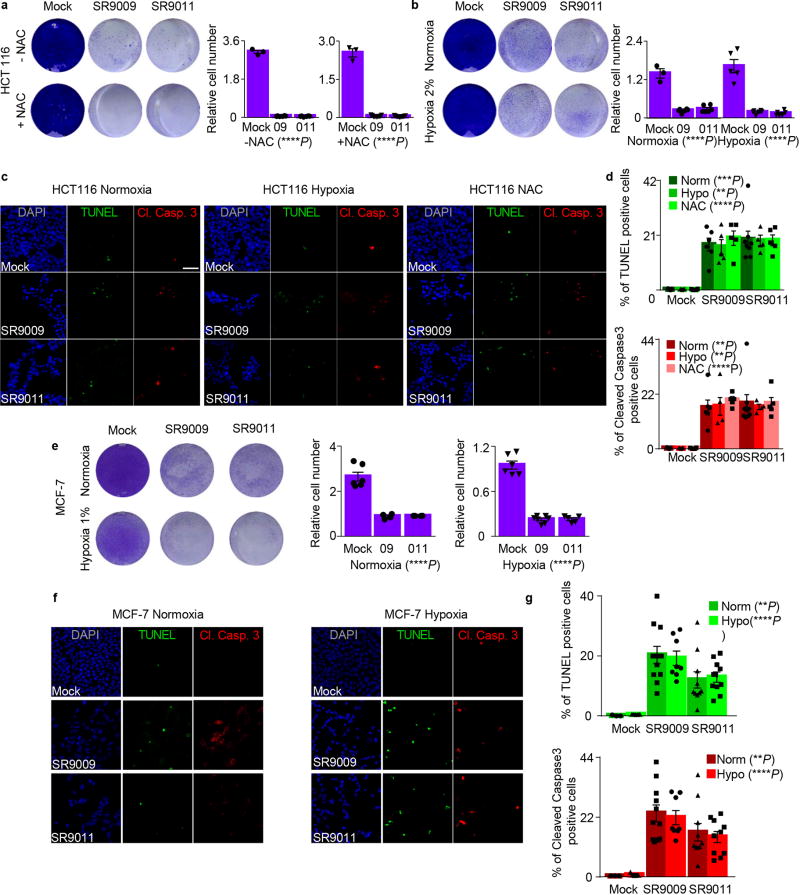
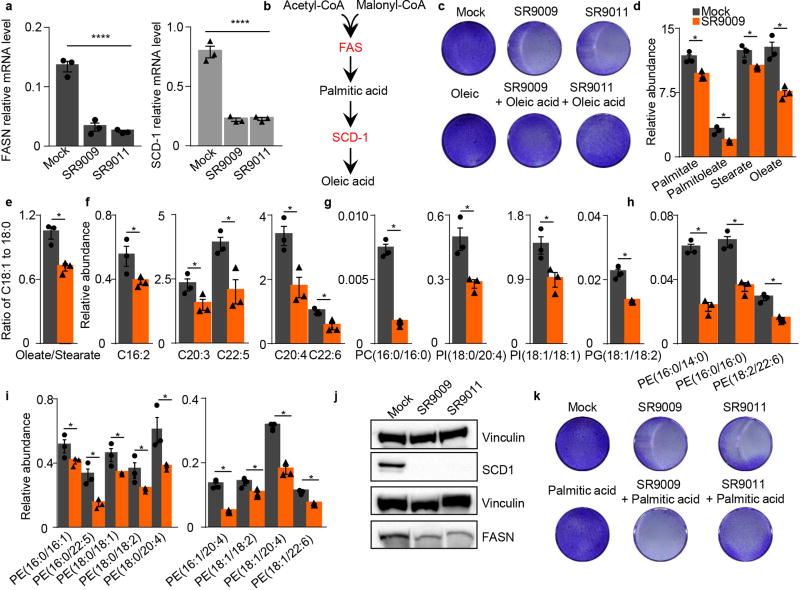
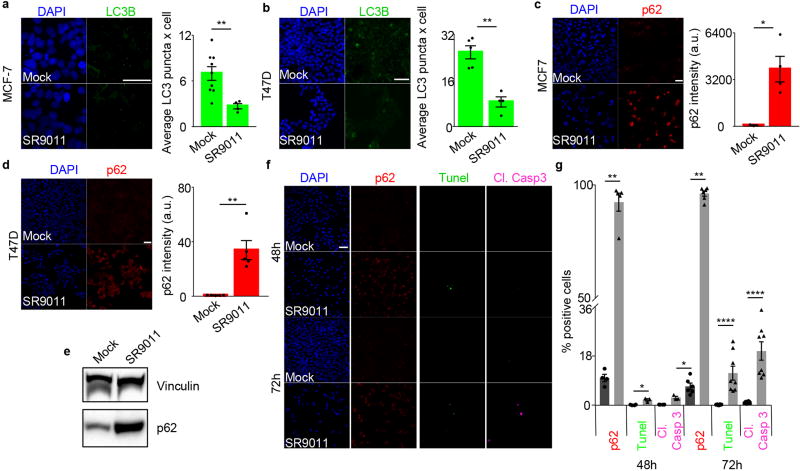
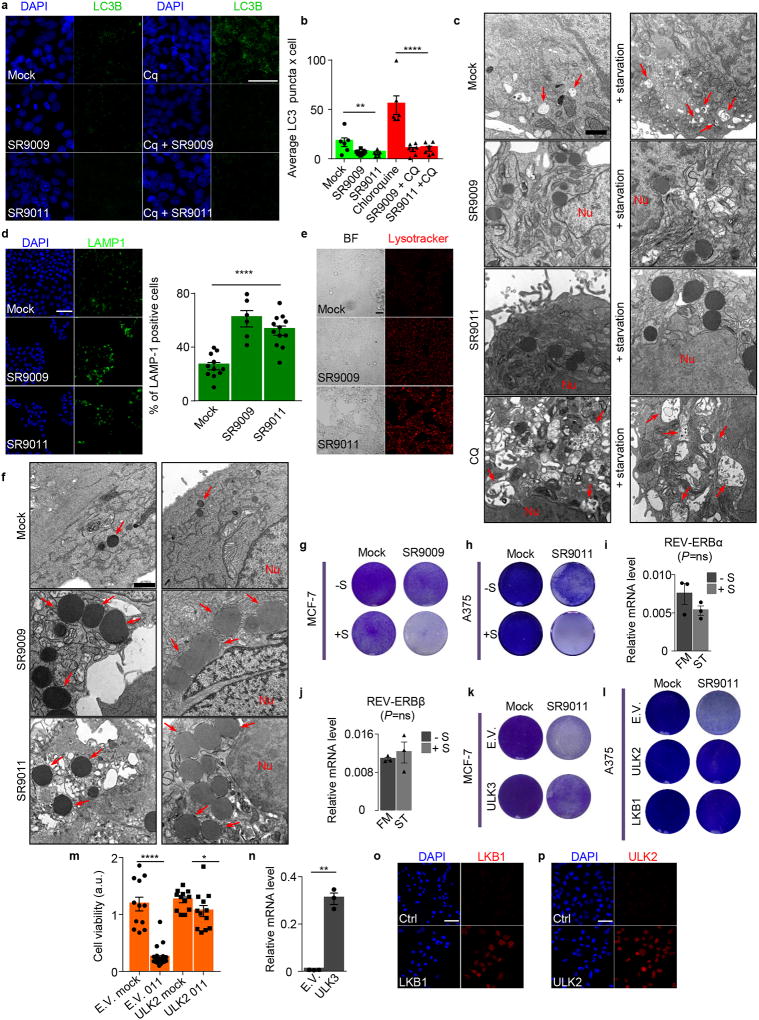
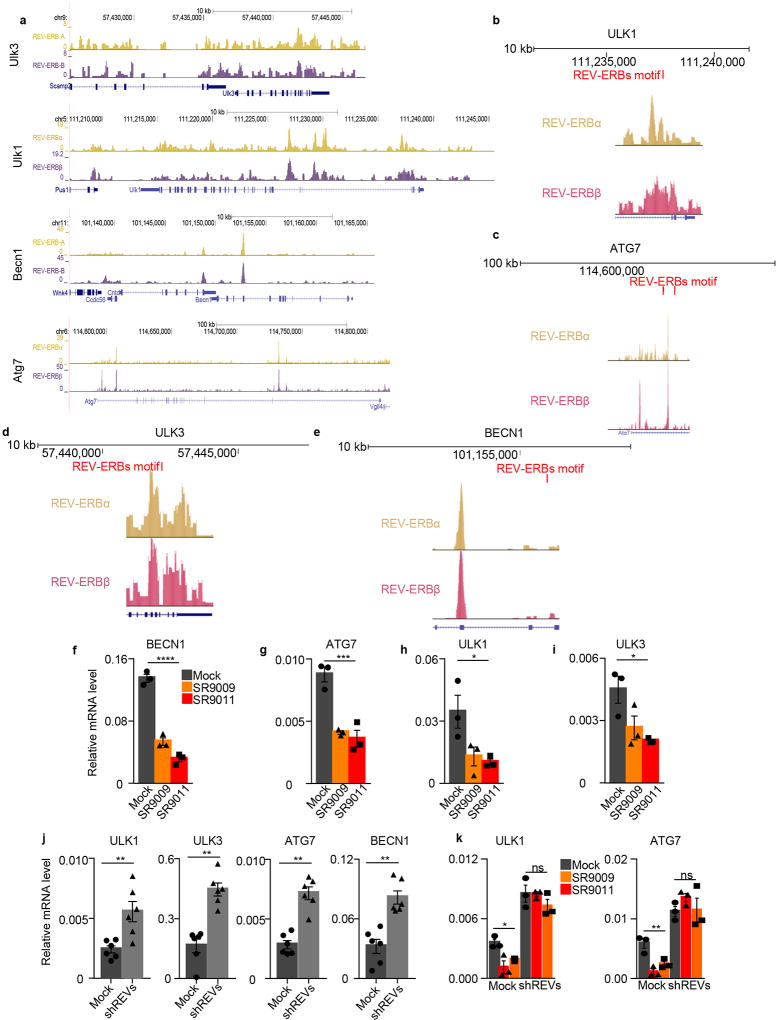
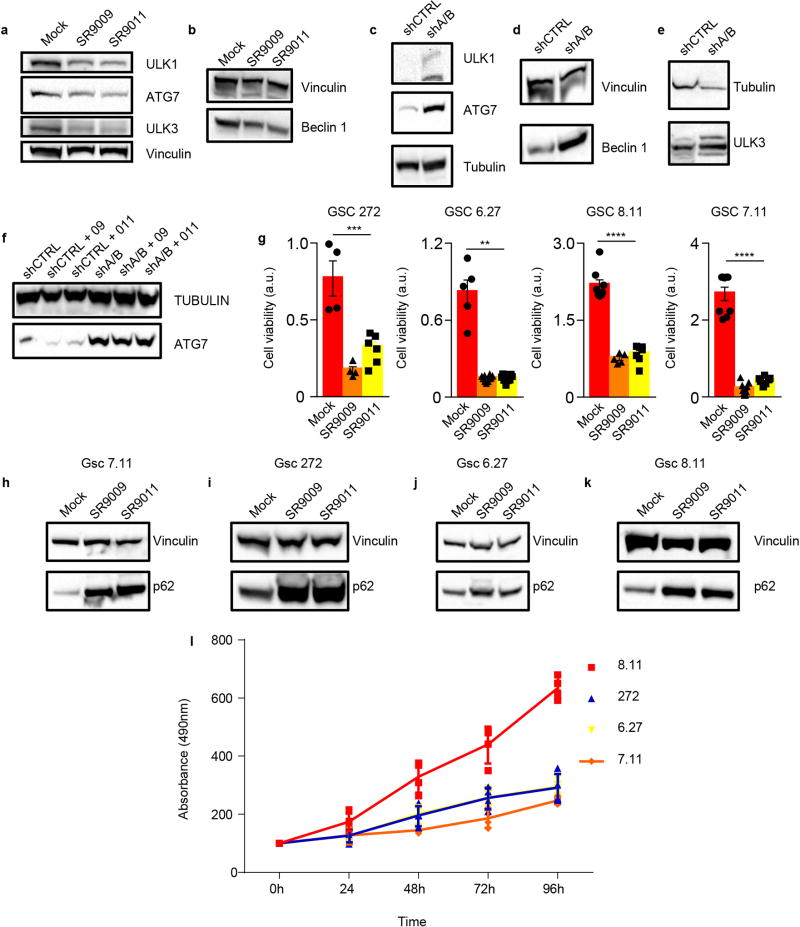
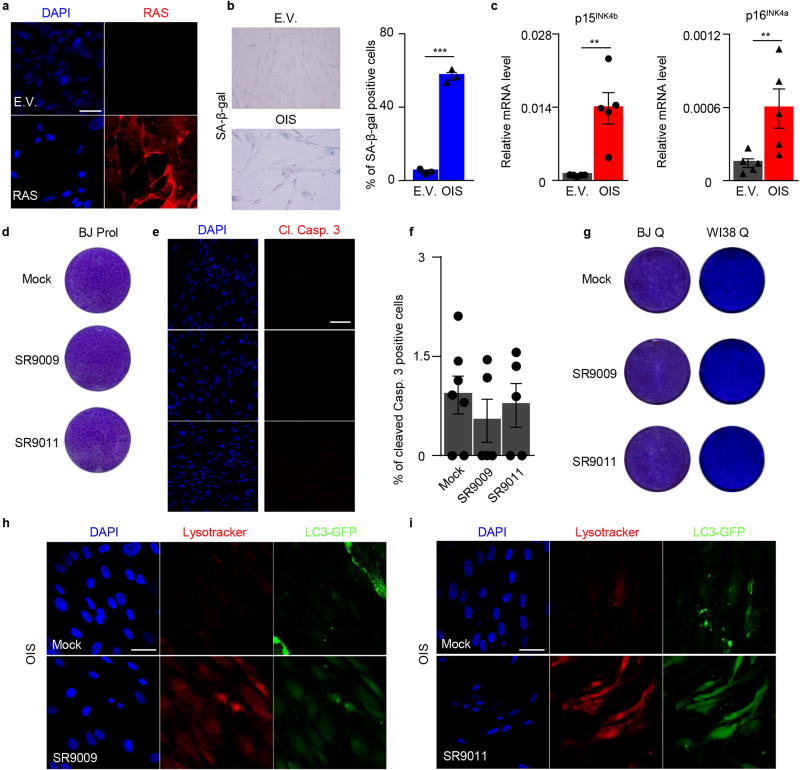
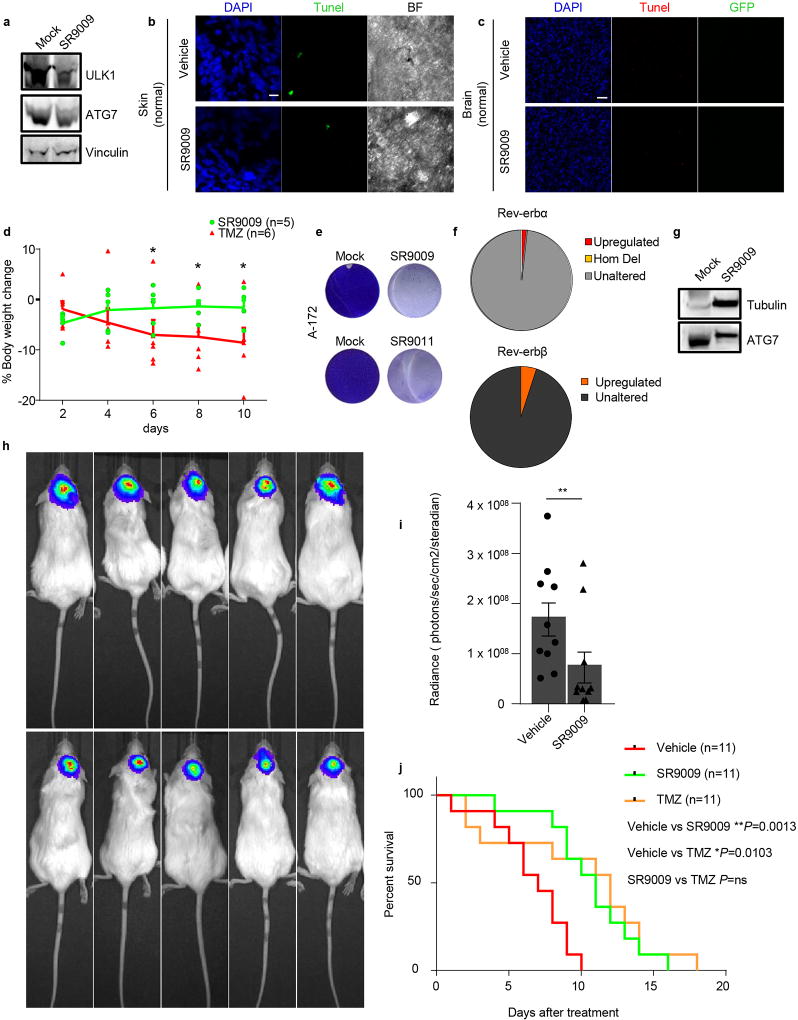
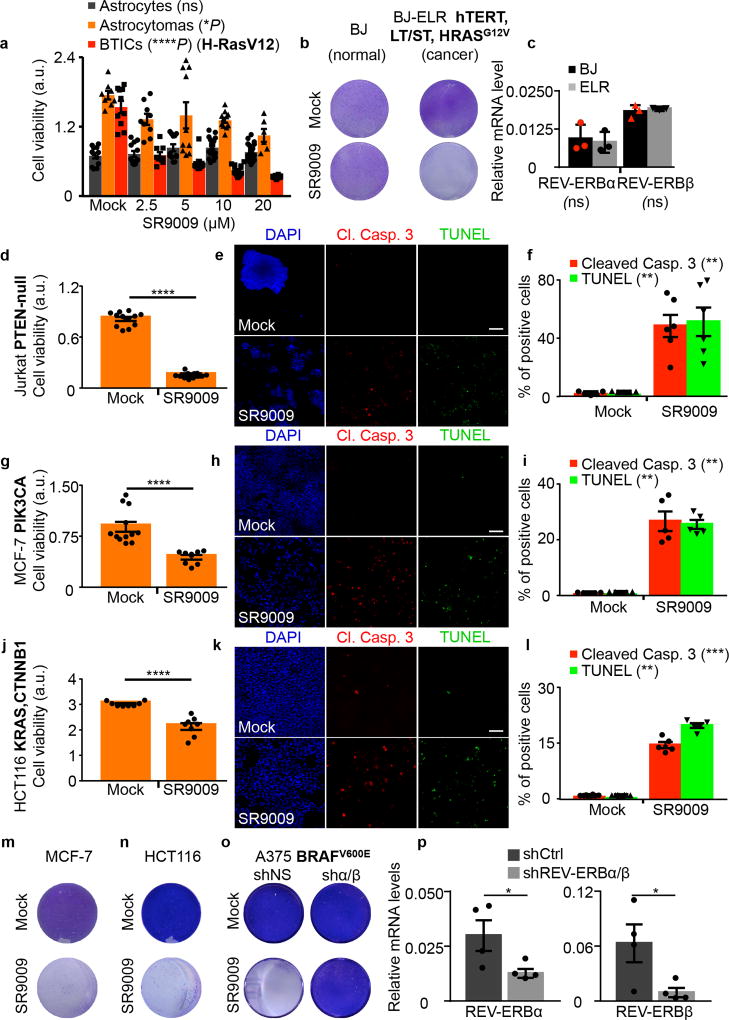
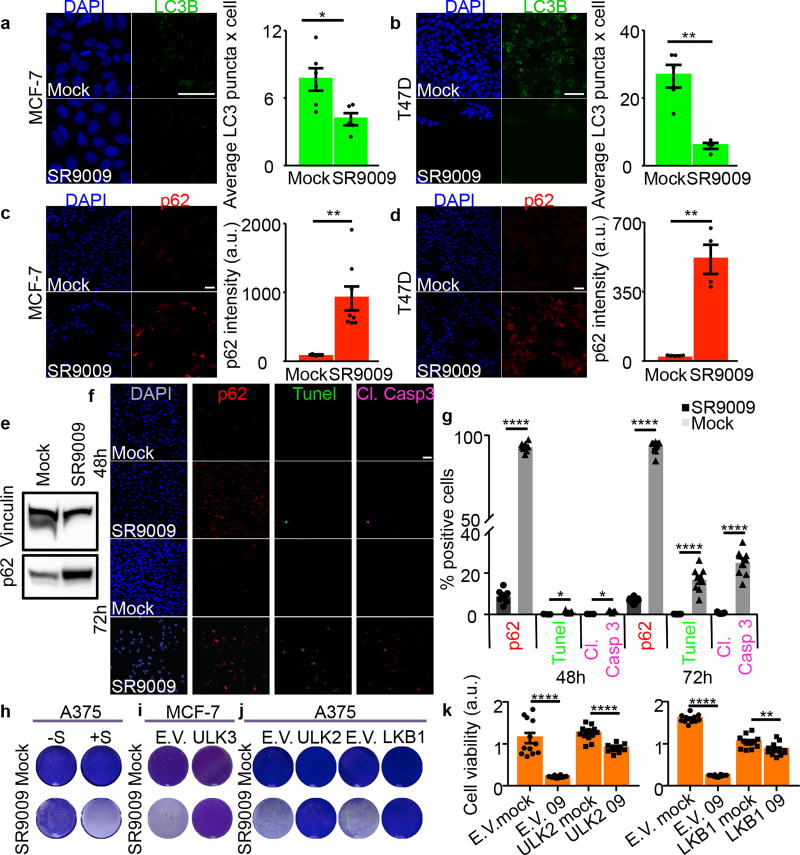
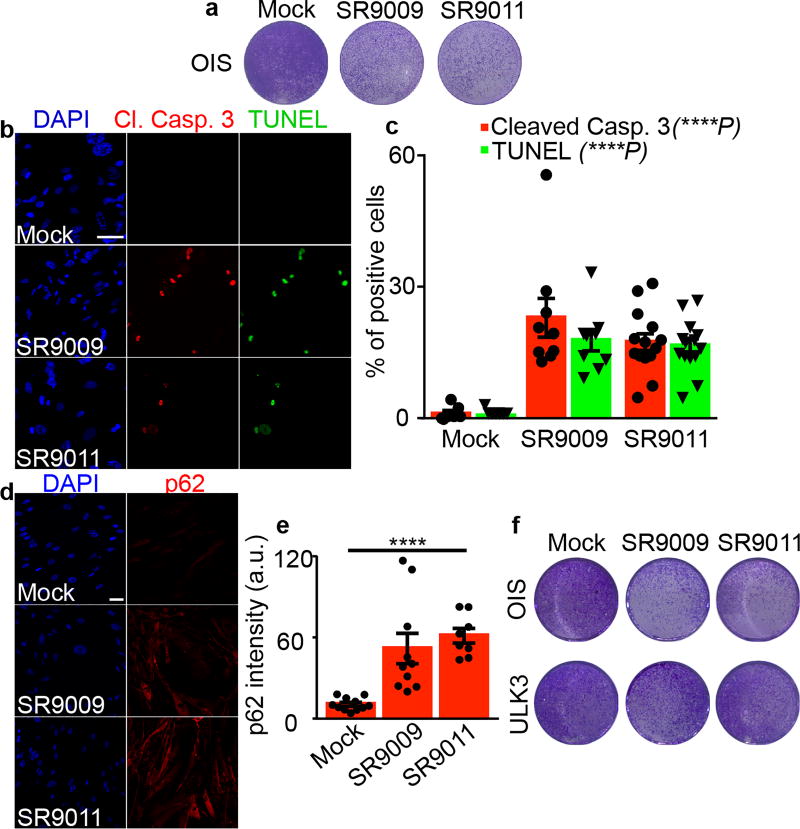
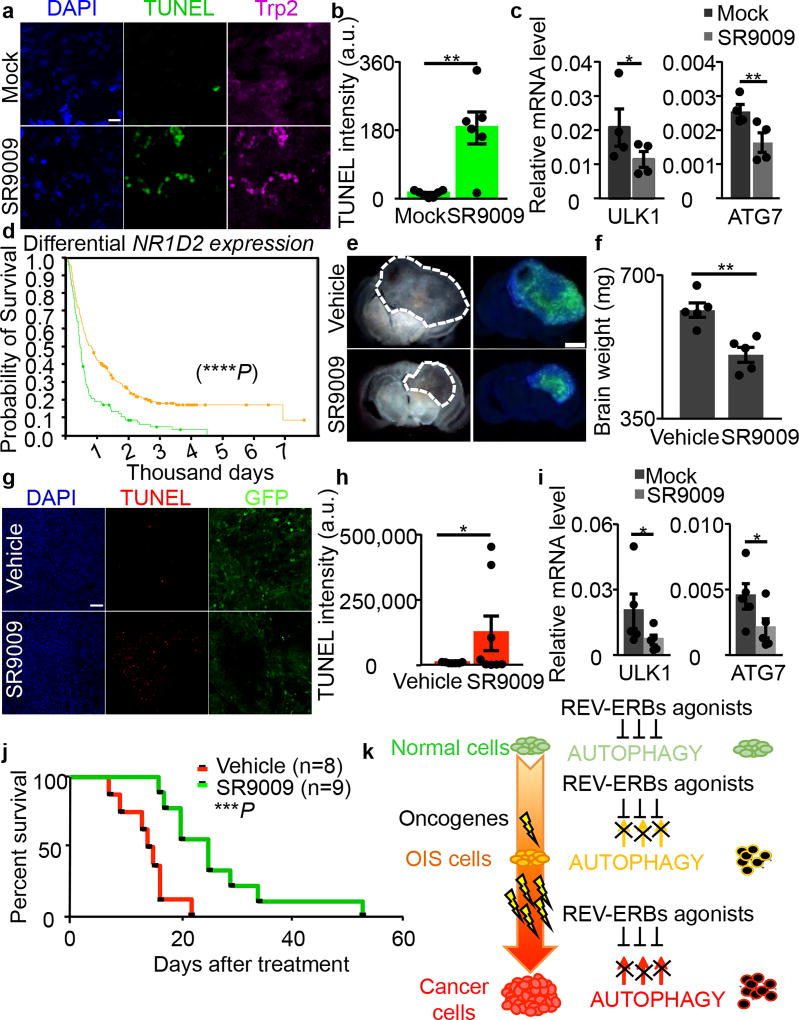
The circadian clock imposes daily rhythms in cell proliferation, metabolism, inflammation and DNA damage response. Perturbations of these processes are hallmarks of cancer and chronic circadian rhythm disruption predisposes individuals to tumour development. This raises the hypothesis that pharmacological modulation of the circadian machinery may be an effective therapeutic strategy for combating cancer. REV-ERBs, the nuclear hormone receptors REV-ERBα (also known as NR1D1) and REV-ERBβ (also known as NR1D2), are essential components of the circadian clock. Here we show that two agonists of REV-ERBs-SR9009 and SR9011-are specifically lethal to cancer cells and oncogene-induced senescent cells, including melanocytic naevi, and have no effect on the viability of normal cells or tissues. The anticancer activity of SR9009 and SR9011 affects a number of oncogenic drivers (such as HRAS, BRAF, PIK3CA and others) and persists in the absence of p53 and under hypoxic conditions. The regulation of autophagy and de novo lipogenesis by SR9009 and SR9011 has a critical role in evoking an apoptotic response in malignant cells. Notably, the selective anticancer properties of these REV-ERB agonists impair glioblastoma growth in vivo and improve survival without causing overt toxicity in mice. These results indicate that pharmacological modulation of circadian regulators is an effective antitumour strategy, identifying a class of anticancer agents with a wide therapeutic window. We propose that REV-ERB agonists are inhibitors of autophagy and de novo lipogenesis, with selective activity towards malignant and benign neoplasms.
Figures
Comment in
-
REV-ERB Agonists Block Autophagy in Cancer Cells.Cancer Discov. 2018 Mar;8(3):261. doi: 10.1158/2159-8290.CD-RW2018-011. Epub 2018 Jan 19. Cancer Discov. 2018. PMID: 29352049
-
Oncogene-induced senescence and tumour control in complex biological systems.Cell Death Differ. 2018 Jun;25(6):1005-1006. doi: 10.1038/s41418-018-0102-y. Epub 2018 Apr 17. Cell Death Differ. 2018. PMID: 29666473 Free PMC article. No abstract available.
References
-
- Fu L, Lee CC. The circadian clock: pacemaker and tumour suppressor. Nature reviews. Cancer. 2003;3:350–361. - PubMed
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. - PubMed
-
- Straif K, et al. Carcinogenicity of shift-work, painting, and fire-fighting. The Lancet. Oncology. 2007;8:1065–1066. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
