

Architecture of a channel-forming O-antigen polysaccharide ABC transporter
- PMID: 29320481
- PMCID: PMC5978415
- DOI: 10.1038/nature25190
Architecture of a channel-forming O-antigen polysaccharide ABC transporter
Abstract
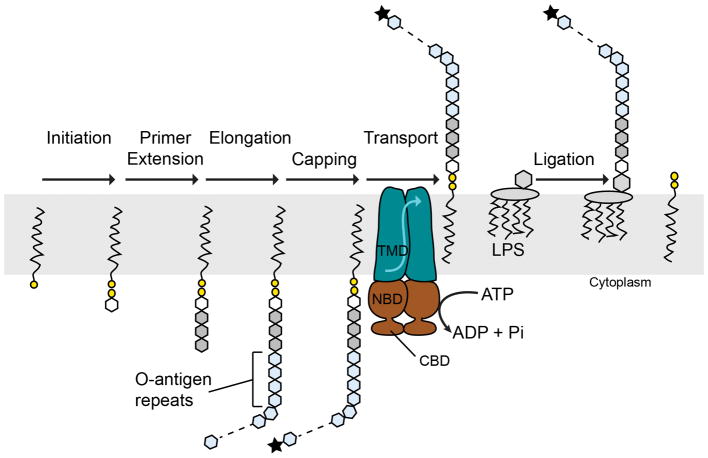
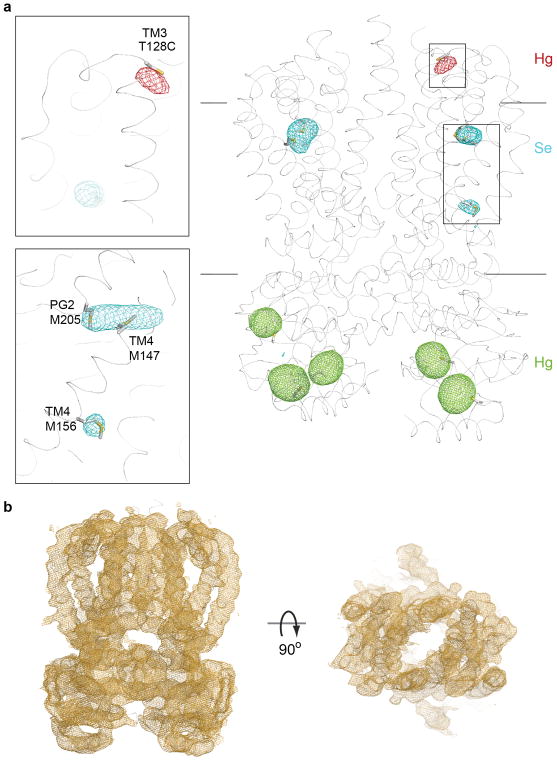
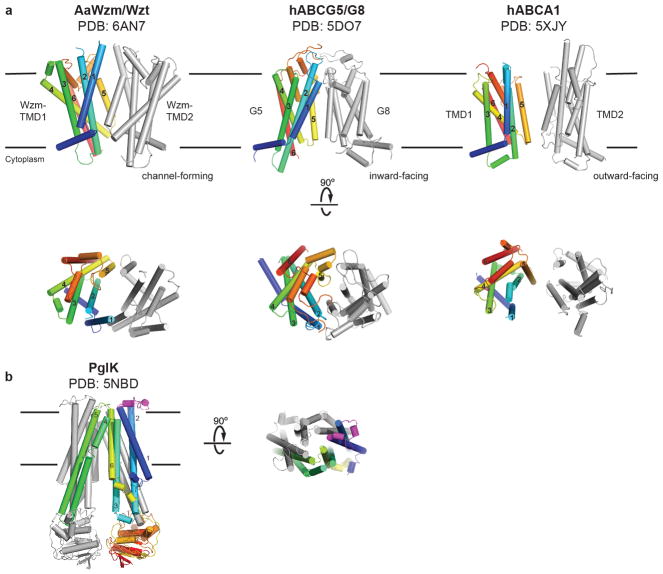
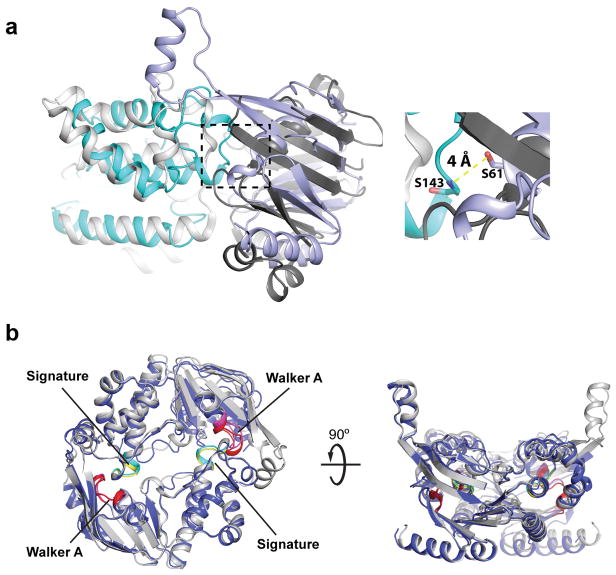
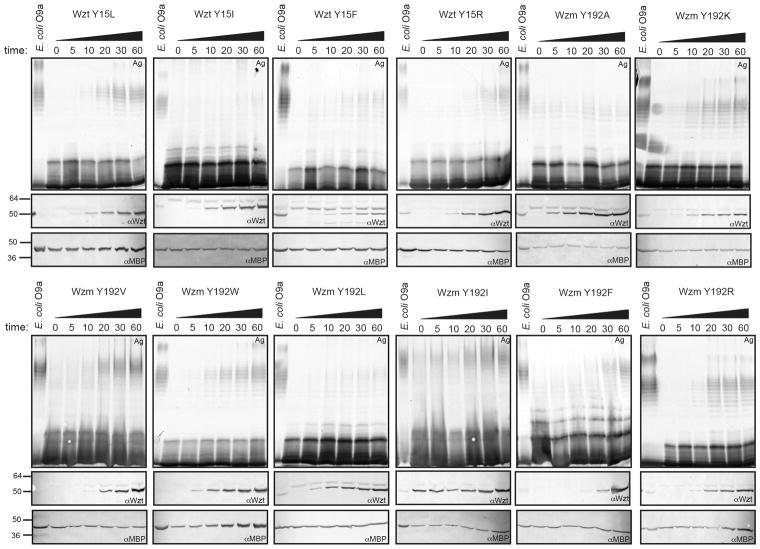
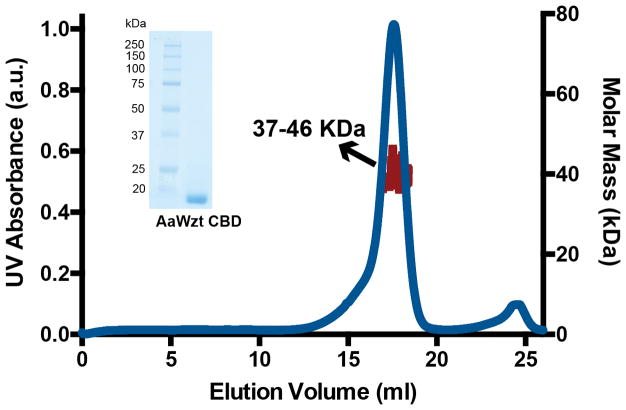
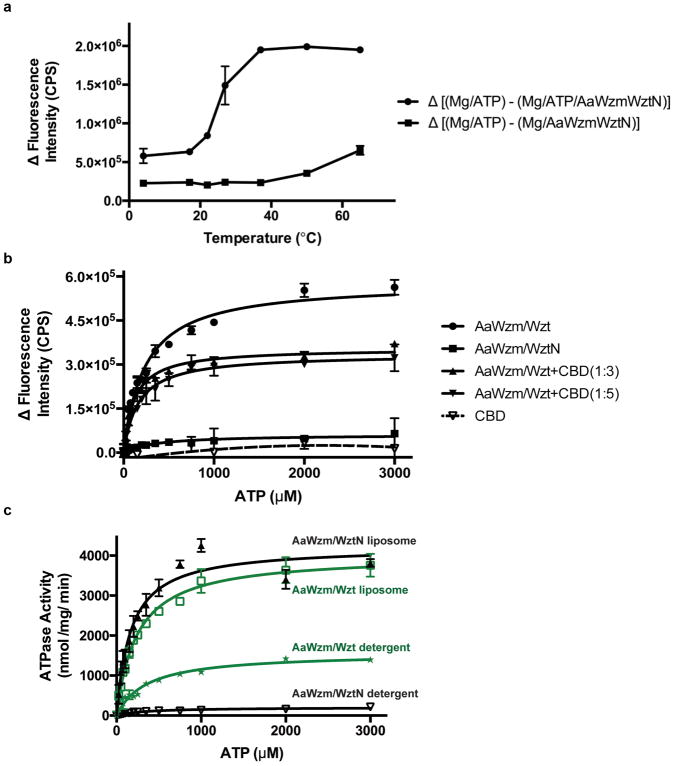
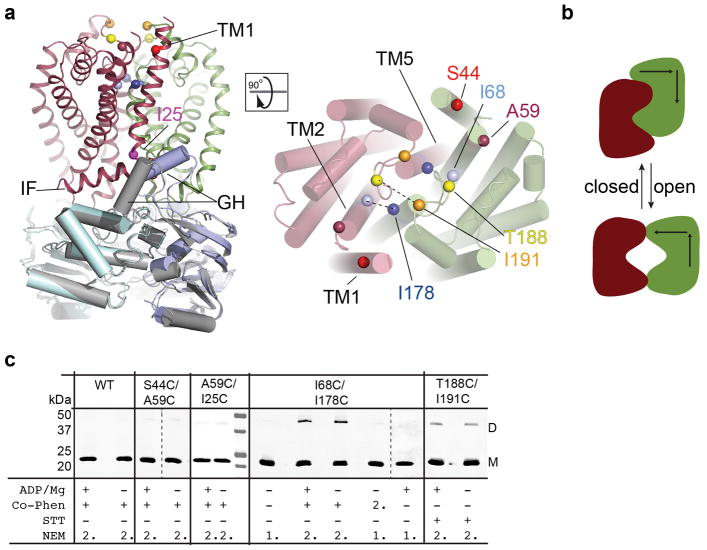
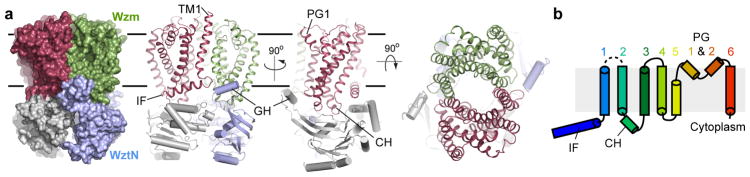
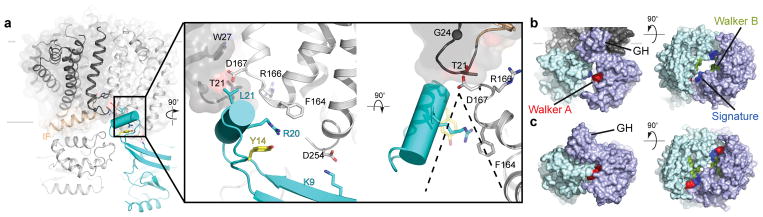
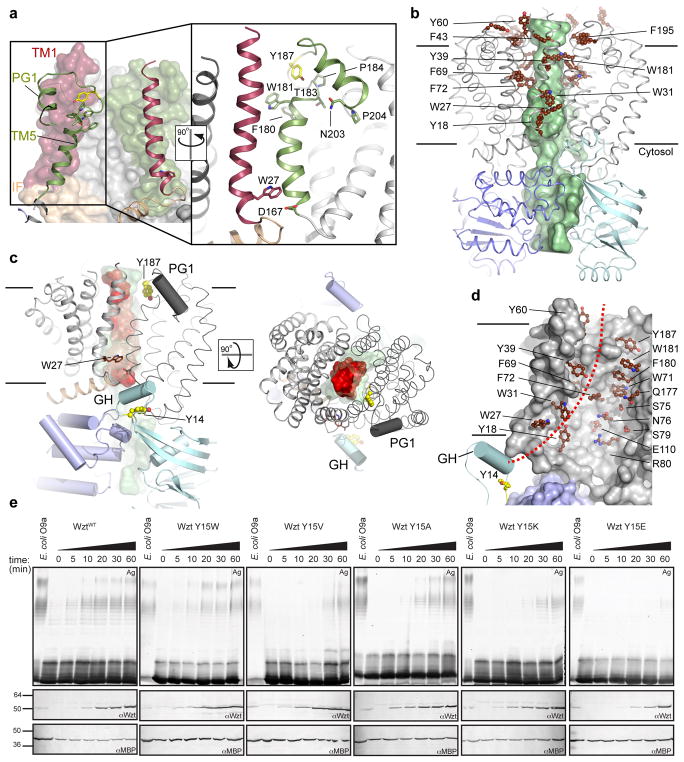
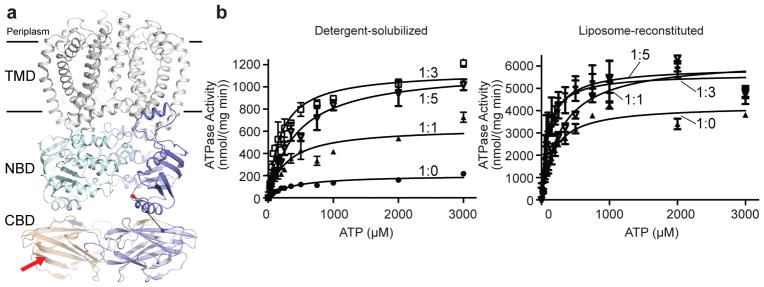
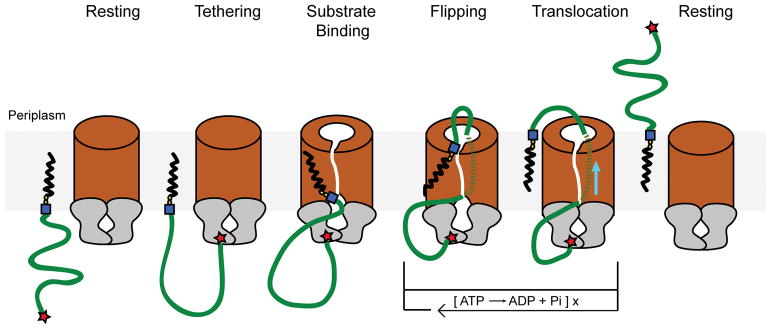
O-antigens are cell surface polysaccharides of many Gram-negative pathogens that aid in escaping innate immune responses. A widespread O-antigen biosynthesis mechanism involves the synthesis of the lipid-anchored polymer on the cytosolic face of the inner membrane, followed by transport to the periplasmic side where it is ligated to the lipid A core to complete a lipopolysaccharide molecule. In this pathway, transport to the periplasm is mediated by an ATP-binding cassette (ABC) transporter, called Wzm-Wzt. Here we present the crystal structure of the Wzm-Wzt homologue from Aquifex aeolicus in an open conformation. The transporter forms a transmembrane channel that is sufficiently wide to accommodate a linear polysaccharide. Its nucleotide-binding domain and a periplasmic extension form 'gate helices' at the cytosolic and periplasmic membrane interfaces that probably serve as substrate entry and exit points. Site-directed mutagenesis of the gates impairs in vivo O-antigen secretion in the Escherichia coli prototype. Combined with a closed structure of the isolated nucleotide-binding domains, our structural and functional analyses suggest a processive O-antigen translocation mechanism, which stands in contrast to the classical alternating access mechanism of ABC transporters.
Conflict of interest statement
The authors declare no competing financial interests
Figures
References
-
- Whitfield C, Szymanski CM, Aebi M. In: Essentials of Glycobiology. Varki A, editor. Chapter 21. 2017. pp. 265–282.
-
- Skurnik M, Bengoechea JA. The biosynthesis and biological role of lipopolysaccharide O-antigens of pathogenic Yersiniae. Carbohydr Res. 2003;338:2521–2529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
