

K-Ras4B Remains Monomeric on Membranes over a Wide Range of Surface Densities and Lipid Compositions
- PMID: 29320680
- PMCID: PMC5984903
- DOI: 10.1016/j.bpj.2017.10.042
K-Ras4B Remains Monomeric on Membranes over a Wide Range of Surface Densities and Lipid Compositions
Abstract
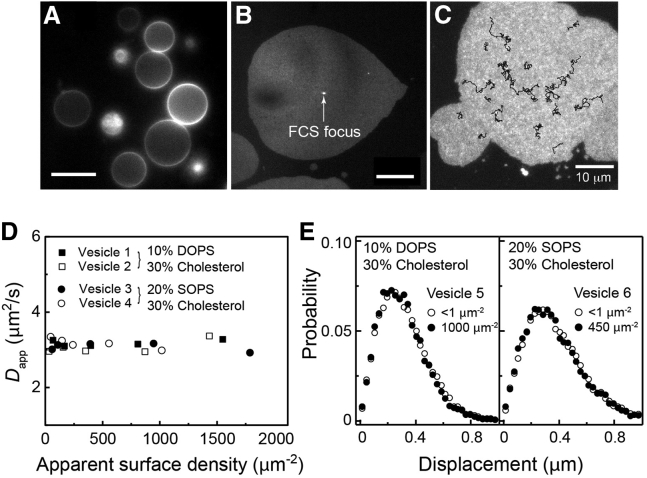
Ras is a membrane-anchored signaling protein that serves as a hub for many signaling pathways and also plays a prominent role in cancer. The intrinsic behavior of Ras on the membrane has captivated the biophysics community in recent years, especially the possibility that it may form dimers. In this article, we describe results from a comprehensive series of experiments using fluorescence correlation spectroscopy and single-molecule tracking to probe the possible dimerization of natively expressed and fully processed K-Ras4B in supported lipid bilayer membranes. Key to these studies is the fact that K-Ras4B has its native membrane anchor, including both the farnesylation and methylation of the terminal cysteine, enabling detailed exploration of possible effects of cholesterol and lipid composition on K-Ras4B membrane organization. The results from all conditions studied indicate that full-length K-Ras4B lacks intrinsic dimerization capability. This suggests that any lateral organization of Ras in living cell membranes likely stems from interactions with other factors.
Copyright © 2017 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
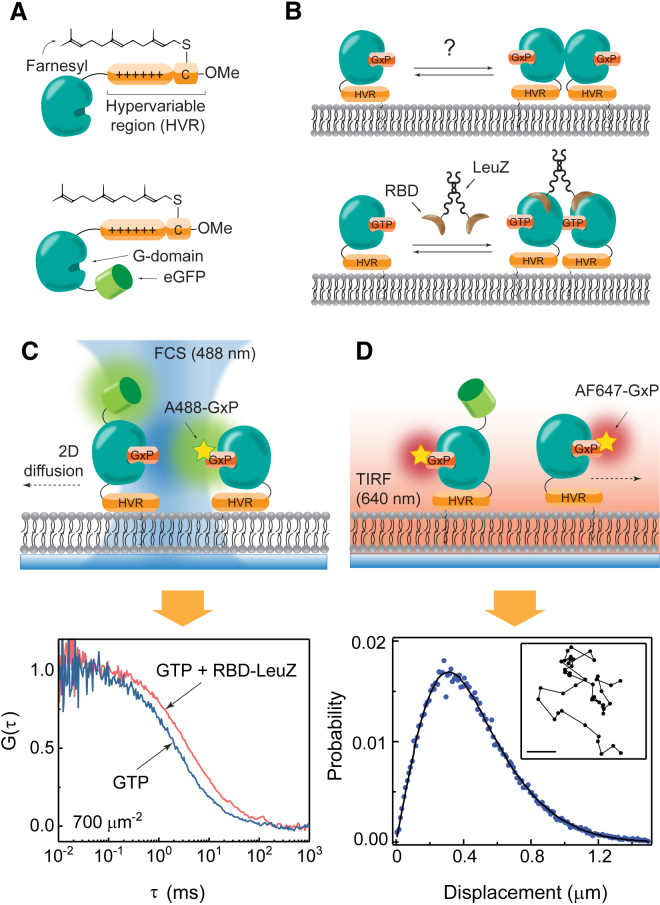
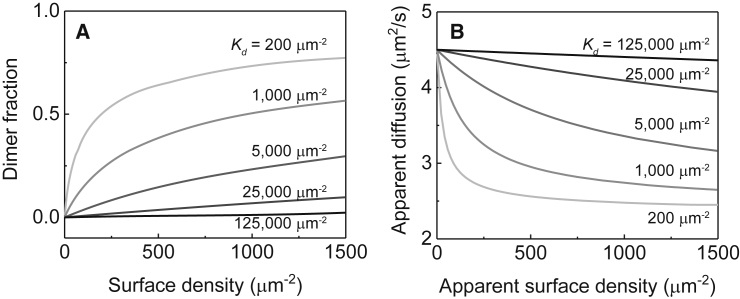
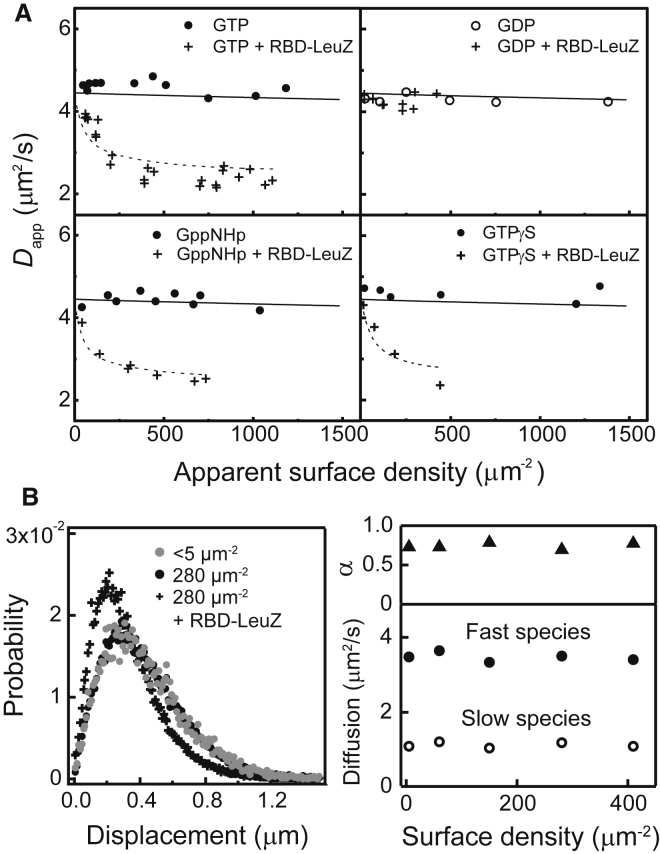
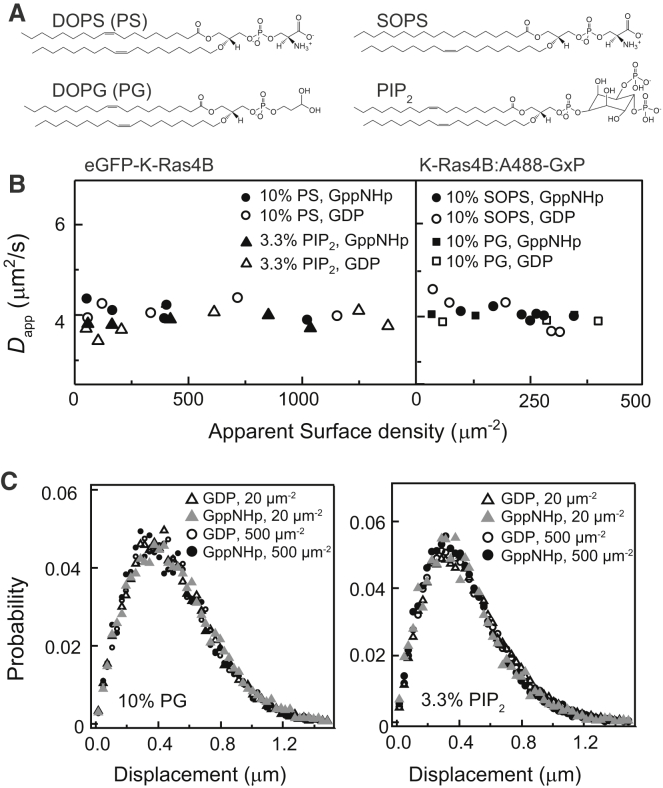
Similar articles
-
Dissociation of the Signaling Protein K-Ras4B from Lipid Membranes Induced by a Molecular Tweezer.Chemistry. 2019 Jul 25;25(42):9827-9833. doi: 10.1002/chem.201901861. Epub 2019 Jul 1. Chemistry. 2019. PMID: 31141233
-
Oncogenic K-Ras4B Dimerization Enhances Downstream Mitogen-activated Protein Kinase Signaling.J Mol Biol. 2020 Feb 14;432(4):1199-1215. doi: 10.1016/j.jmb.2020.01.002. Epub 2020 Jan 10. J Mol Biol. 2020. PMID: 31931009 Free PMC article.
-
Mechanisms of membrane binding of small GTPase K-Ras4B farnesylated hypervariable region.J Biol Chem. 2015 Apr 10;290(15):9465-77. doi: 10.1074/jbc.M114.620724. Epub 2015 Feb 24. J Biol Chem. 2015. PMID: 25713064 Free PMC article.
-
The Hypervariable Region of K-Ras4B Governs Molecular Recognition and Function.Int J Mol Sci. 2019 Nov 14;20(22):5718. doi: 10.3390/ijms20225718. Int J Mol Sci. 2019. PMID: 31739603 Free PMC article. Review.
-
Diffusion-based determination of protein homodimerization on reconstituted membrane surfaces.BMB Rep. 2021 Mar;54(3):157-163. doi: 10.5483/BMBRep.2021.54.3.228. BMB Rep. 2021. PMID: 33408000 Free PMC article. Review.
Cited by
-
Biology, pathology, and therapeutic targeting of RAS.Adv Cancer Res. 2020;148:69-146. doi: 10.1016/bs.acr.2020.05.002. Epub 2020 Jul 9. Adv Cancer Res. 2020. PMID: 32723567 Free PMC article. Review.
-
The Ras dimer structure.Chem Sci. 2021 May 4;12(23):8178-8189. doi: 10.1039/d1sc00957e. Chem Sci. 2021. PMID: 34194708 Free PMC article.
-
Membrane lipids drive formation of KRAS4b-RAF1 RBDCRD nanoclusters on the membrane.Commun Biol. 2024 Feb 28;7(1):242. doi: 10.1038/s42003-024-05916-0. Commun Biol. 2024. PMID: 38418613 Free PMC article.
-
Machine learning-driven multiscale modeling reveals lipid-dependent dynamics of RAS signaling proteins.Proc Natl Acad Sci U S A. 2022 Jan 4;119(1):e2113297119. doi: 10.1073/pnas.2113297119. Proc Natl Acad Sci U S A. 2022. PMID: 34983849 Free PMC article.
-
RAS Function in cancer cells: translating membrane biology and biochemistry into new therapeutics.Biochem J. 2020 Aug 14;477(15):2893-2919. doi: 10.1042/BCJ20190839. Biochem J. 2020. PMID: 32797215 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
