

DNA Unwinding Is the Primary Determinant of CRISPR-Cas9 Activity
- PMID: 29320733
- PMCID: PMC11151164
- DOI: 10.1016/j.celrep.2017.12.041
DNA Unwinding Is the Primary Determinant of CRISPR-Cas9 Activity
Abstract
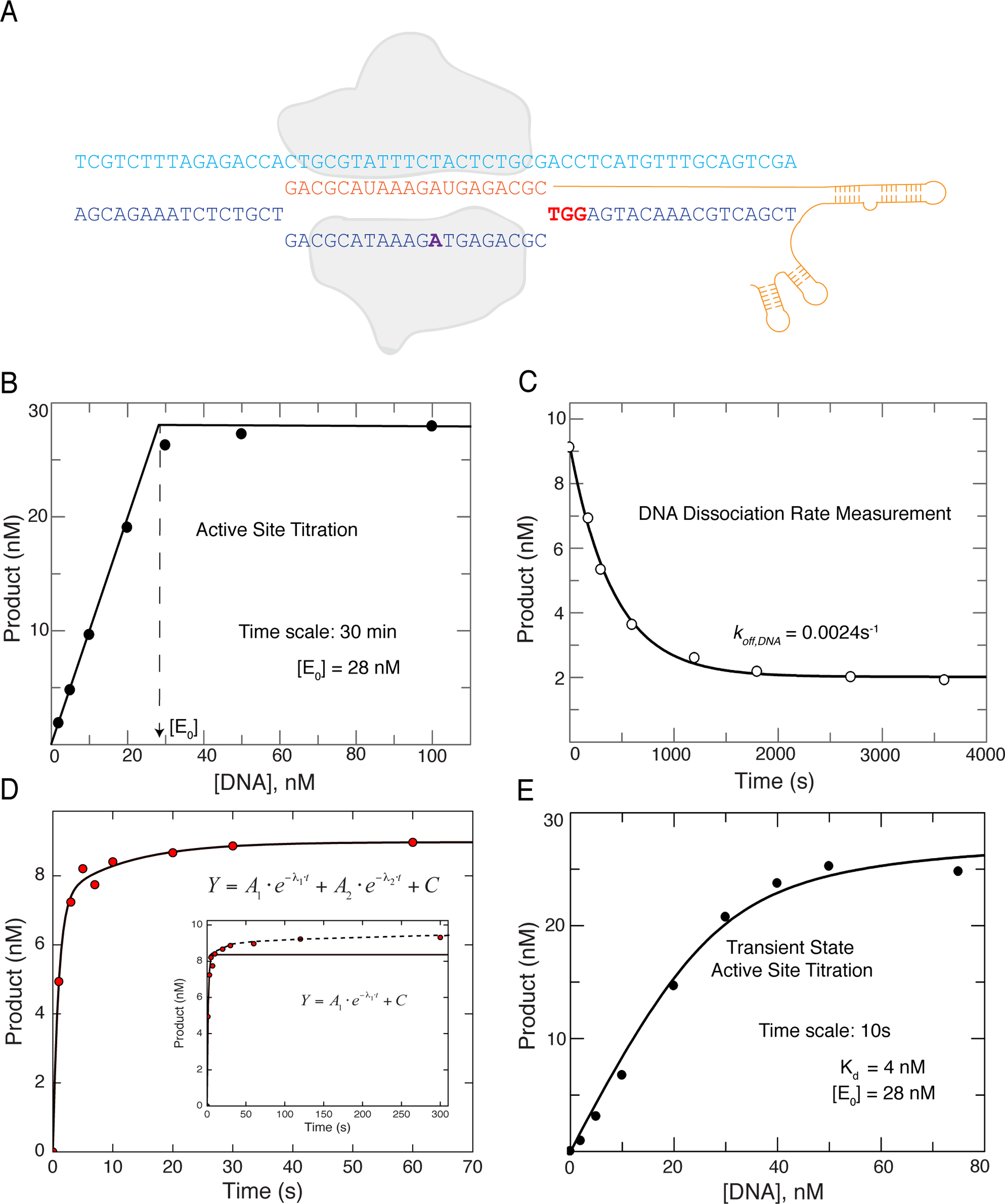
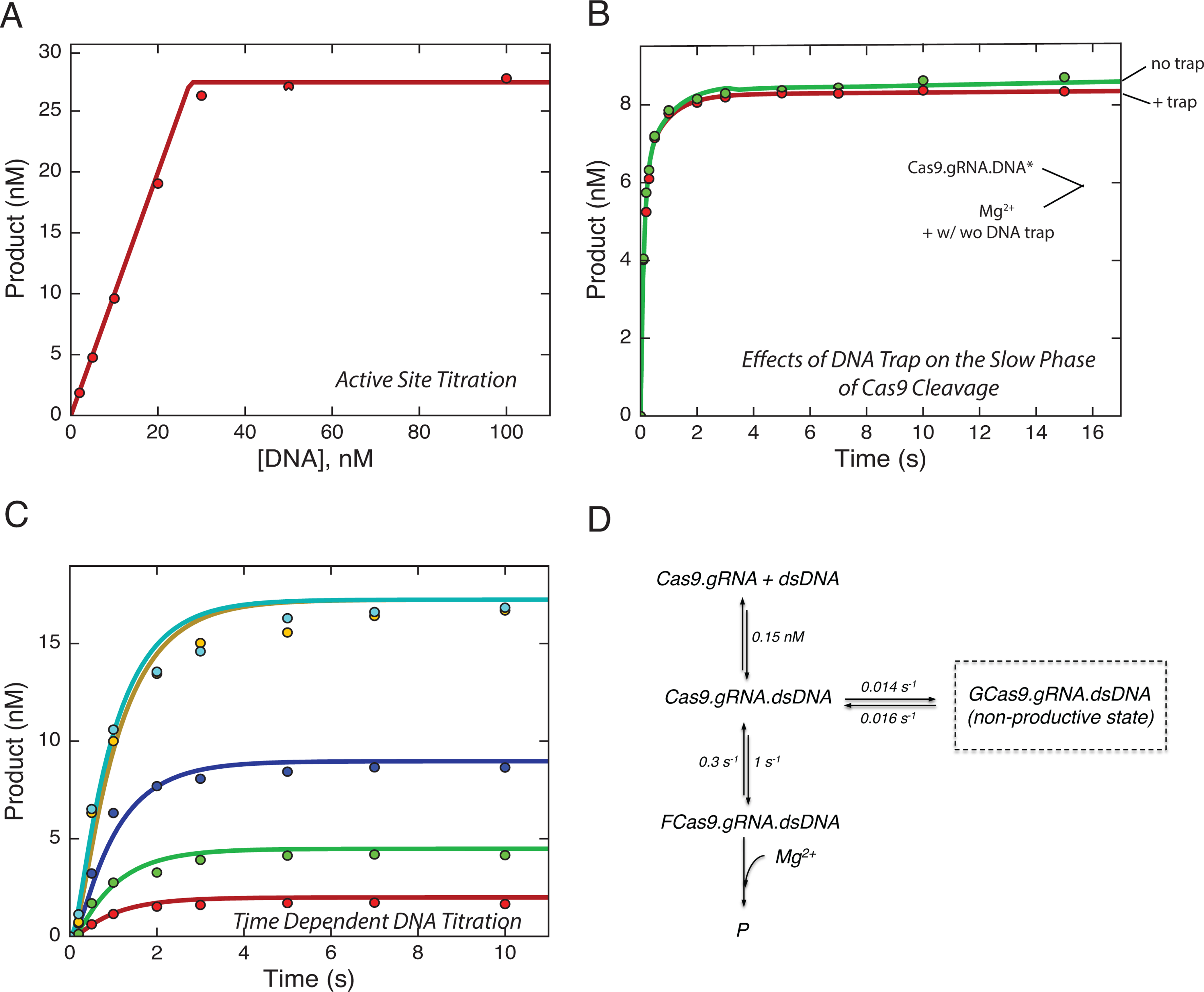
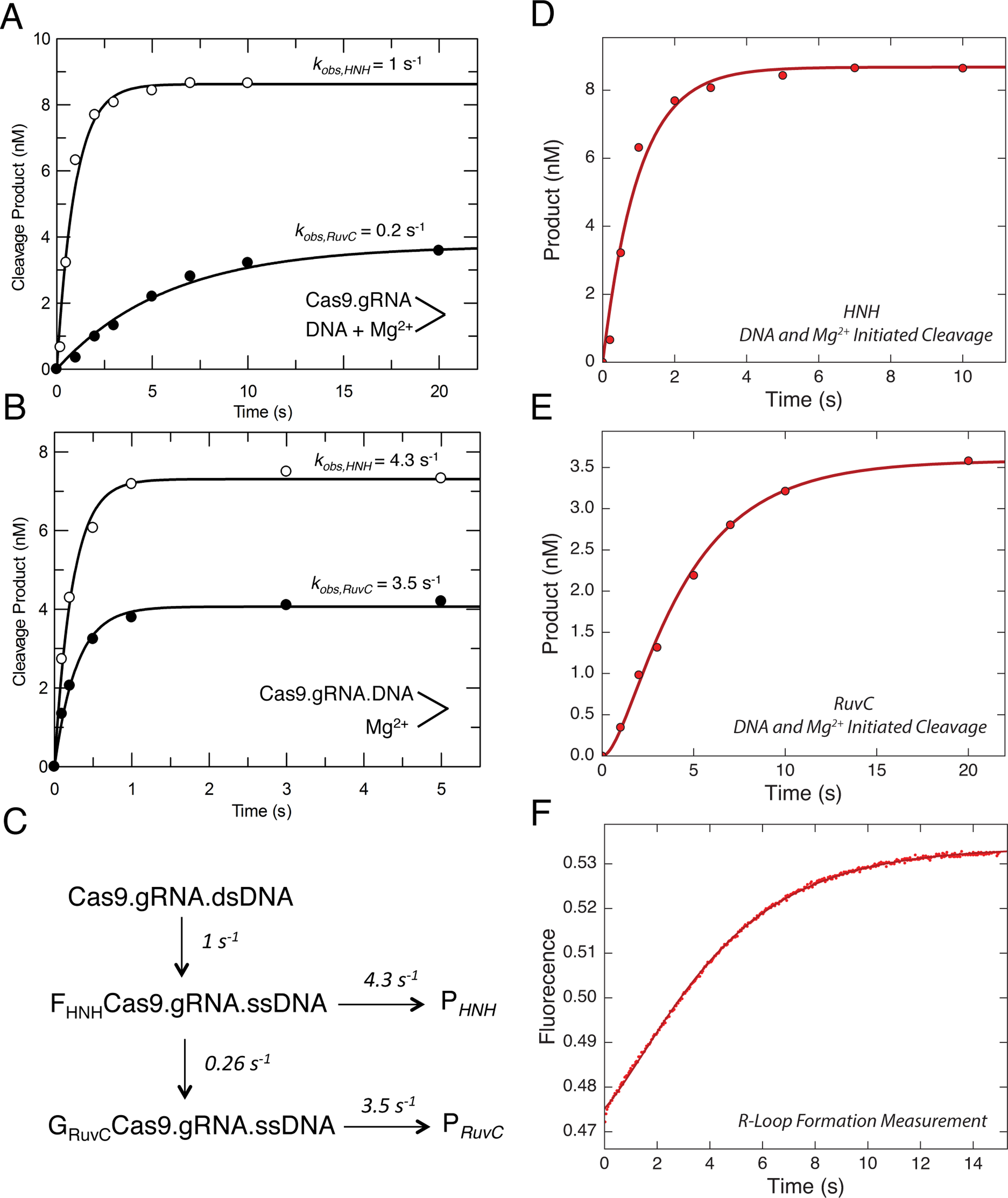
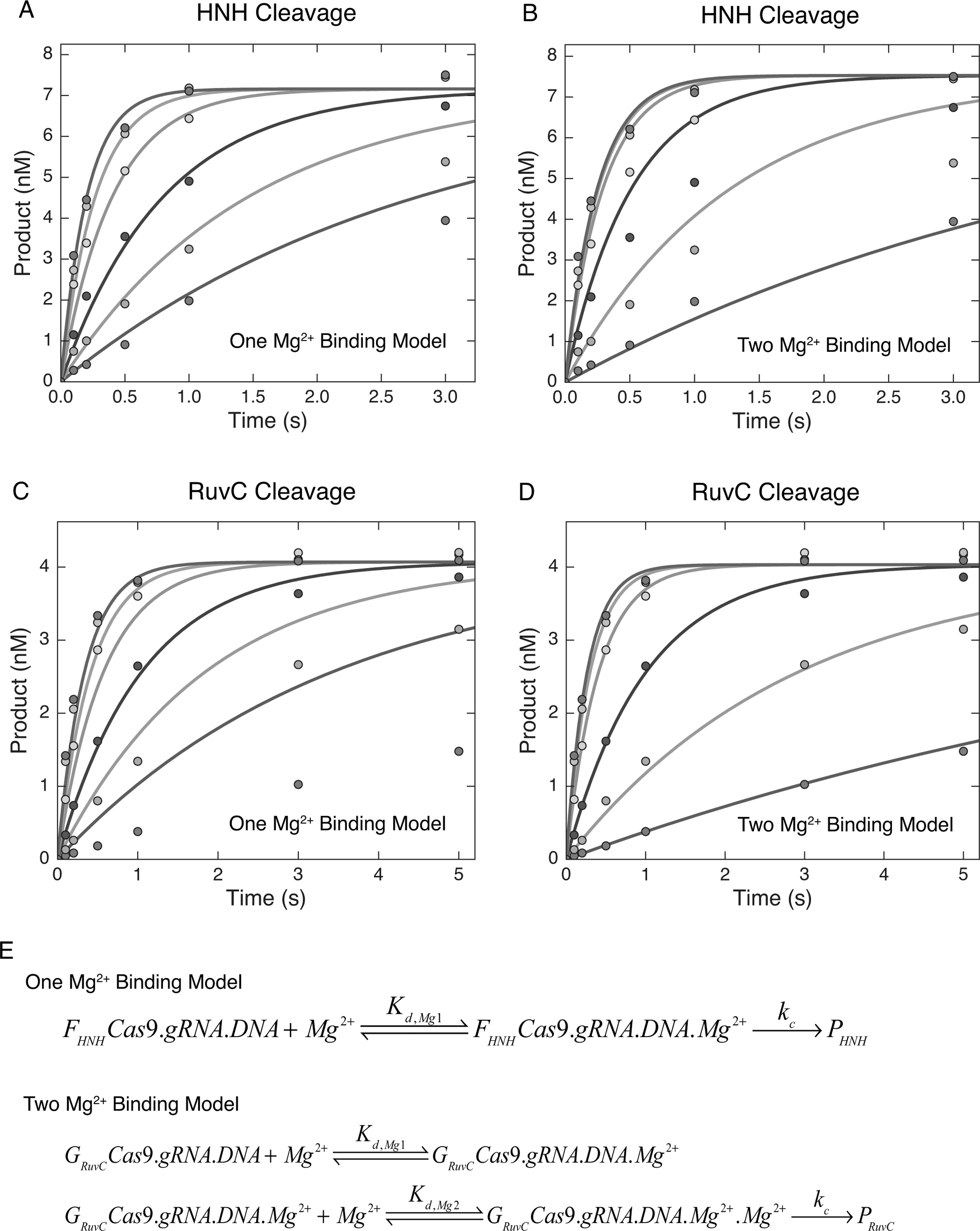
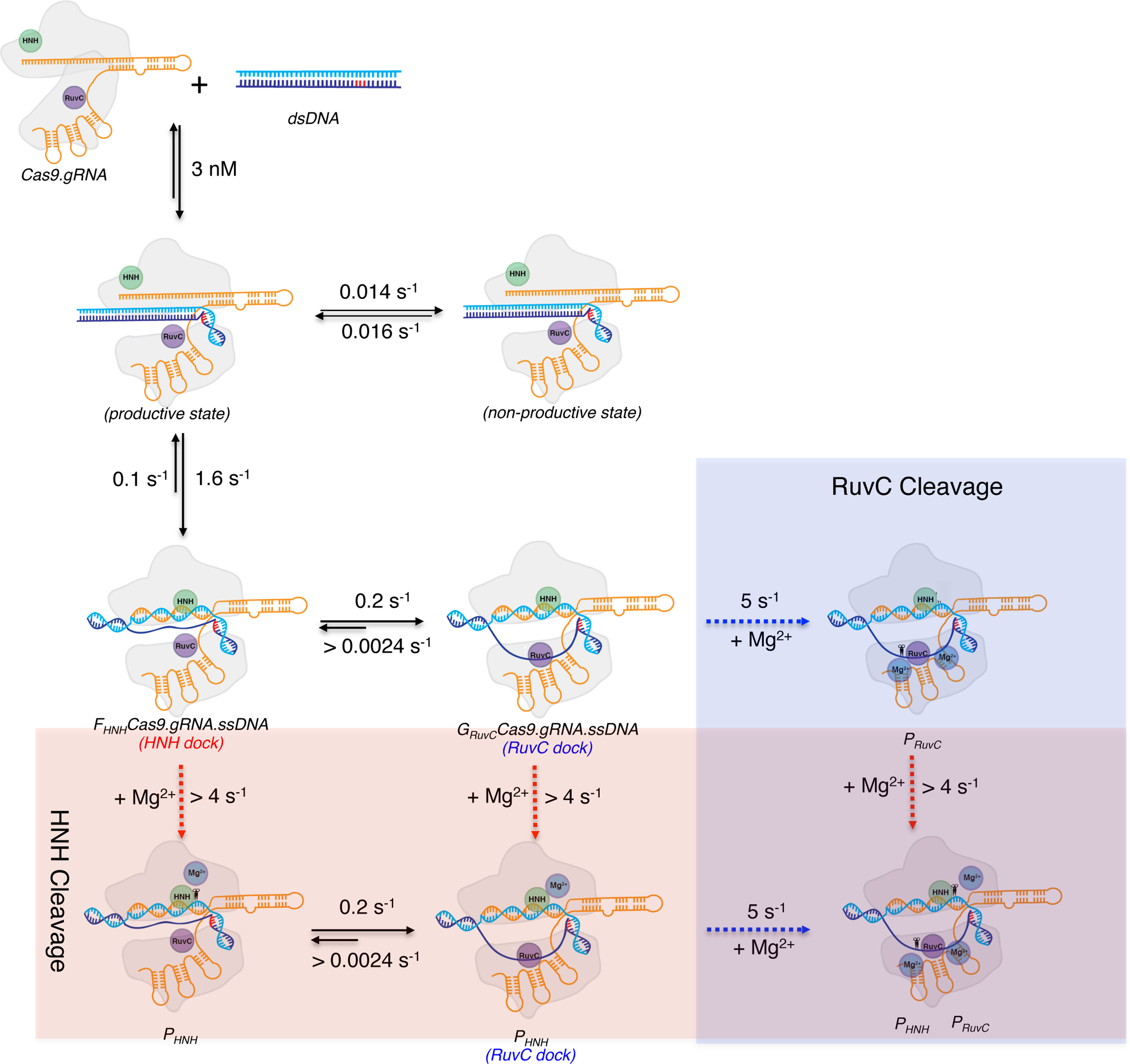
Bacterial adaptive immunity utilizes RNA-guided surveillance complexes comprising Cas proteins together with CRISPR RNAs (crRNAs) to target foreign nucleic acids for destruction. Cas9, a type II CRISPR-Cas effector complex, can be programed with a single-guide RNA that base pairs with the target strand of dsDNA, displacing the non-target strand to create an R-loop, where the HNH and the RuvC nuclease domains cleave opposing strands. While many structural and biochemical studies have shed light on the mechanism of Cas9 cleavage, a clear unifying model has yet to emerge. Our detailed kinetic characterization of the enzyme reveals that DNA binding is reversible, and R-loop formation is rate-limiting, occurring in two steps, one for each of the nuclease domains. The specificity constant for cleavage is determined through an induced-fit mechanism as the product of the equilibrium binding affinity for DNA and the rate of R-loop formation.
Keywords: CRISPR; Cas9; DNA cleavage; R-loop; kinetics; sgRNA.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Amitai G. and Sorek R. (2016) ‘CRISPR-Cas adaptation: insights into the mechanism of action’, Nat Rev Microbiol, 14(2), pp. 67–76. - PubMed
-
- Ariyoshi M, Vassylyev DG, Iwasaki H, Nakamura H, Shinagawa H. and Morikawa K. (1994) ‘Atomic structure of the RuvC resolvase: a holliday junction-specific endonuclease from E. coli’, Cell, 78(6), pp. 1063–72. - PubMed
-
- Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, Romero DA and Horvath P. (2007) ‘CRISPR provides acquired resistance against viruses in prokaryotes’, Science, 315(5819), pp. 1709–12. - PubMed
-
- Biertümpfel C, Yang W. and Suck D. (2007) ‘Crystal structure of T4 endonuclease VII resolving a Holliday junction’, Nature, 449(7162), pp. 616–20. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
