

Dual action of highbush blueberry proanthocyanidins on Aggregatibacter actinomycetemcomitans and the host inflammatory response
- PMID: 29321009
- PMCID: PMC5763534
- DOI: 10.1186/s12906-017-2072-x
Dual action of highbush blueberry proanthocyanidins on Aggregatibacter actinomycetemcomitans and the host inflammatory response
Abstract
Background: The highbush blueberry (Vaccinium corymbosum) has a beneficial effect on several aspects of human health. The present study investigated the effects of highbush blueberry proanthocyanidins (PACs) on the virulence properties of Aggregatibacter actinomycetemcomitans and macrophage-associated inflammatory responses.
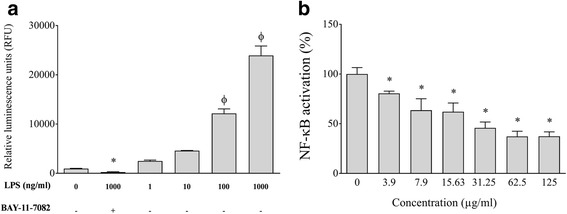
Methods: PACs were isolated from frozen highbush blueberries using solid-phase chromatography. A microplate dilution assay was performed to determine the effect of highbush blueberry PACs on A. actinomycetemcomitans growth as well as biofilm formation stained with crystal violet. Tight junction integrity of oral keratinocytes was assessed by measuring the transepithelial electrical resistance (TER), while macrophage viability was determined with a colorimetric MTT assay. Pro-inflammatory cytokine and MMP secretion by A. actinomycetemcomitans-stimulated macrophages was quantified by ELISA. The U937-3xκB-LUC monocyte cell line transfected with a luciferase reporter gene was used to monitor NF-κB activation.
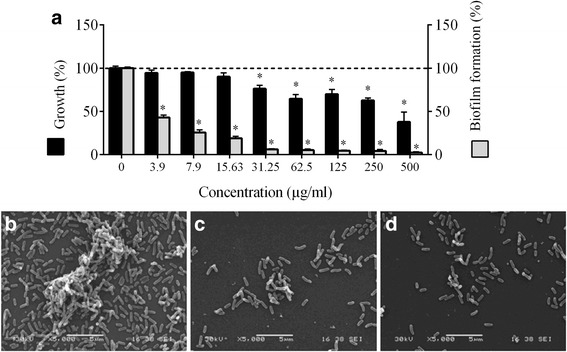
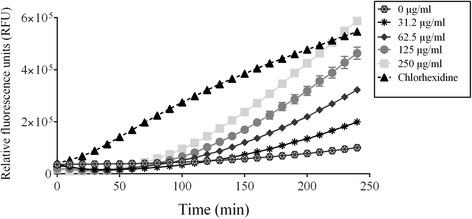
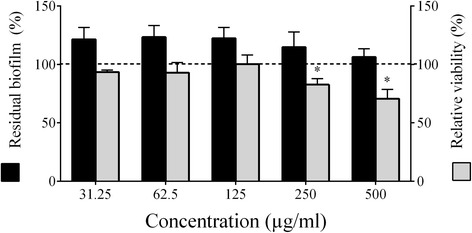
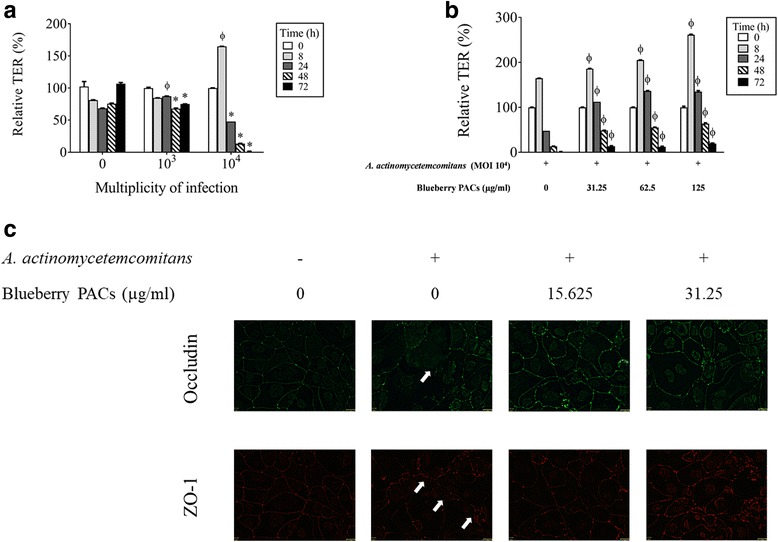
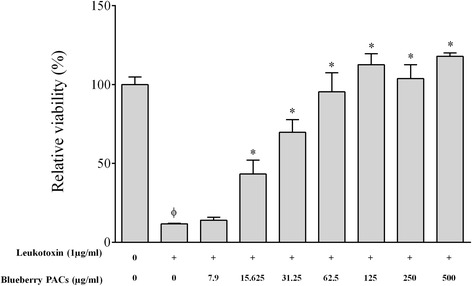
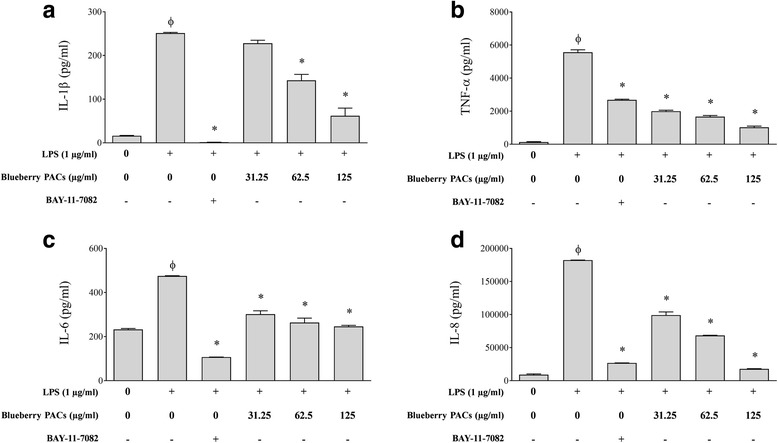
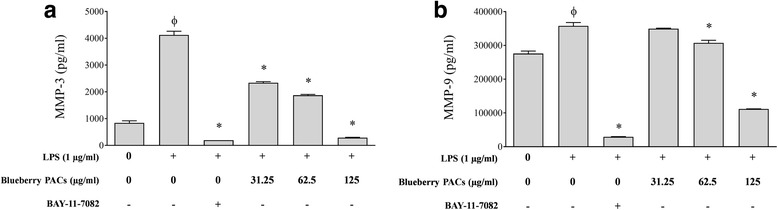
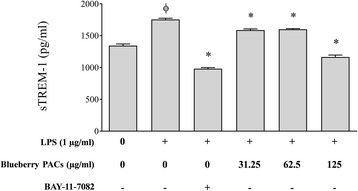
Results: Highbush blueberry PACs reduced the growth of A. actinomycetemcomitans and prevented biofilm formation at sub-inhibitory concentrations. The treatment of pre-formed biofilms with the PACs resulted in a loss of bacterial viability. The antibacterial activity of the PACs appeared to involve damage to the bacterial cell membrane. The PACs protected the oral keratinocytes barrier integrity from damage caused by A. actinomycetemcomitans. The PACs also protected macrophages from the deleterious effect of leukotoxin Ltx-A and dose-dependently inhibited the secretion of pro-inflammatory cytokines (IL-1β, IL-6, CXCL8, TNF-α), matrix metalloproteinases (MMP-3, MMP-9), and sTREM-1 by A. actinomycetemcomitans-treated macrophages. The PACs also inhibited the activation of the NF-κB signaling pathway.
Conclusion: The antibacterial and anti-inflammatory properties of highbush blueberry PACs as well as their ability to protect the oral keratinocyte barrier and neutralize leukotoxin activity suggest that they may be promising candidates as novel therapeutic agents.
Keywords: Aggregatibacter actinomycetemcomitans; Blueberry; Cytokine; Keratinocyte; Leukotoxin; Macrophages; Matrix metalloproteinase; Periodontal disease; Proanthocyanidins; Tight junction.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Socransky SS, Haffajee AD. The bacterial etiology of destructive periodontal disease: current concepts. J Periodontol. 1992;63(4 Suppl):322–331. - PubMed
-
- Fine DH, Markowitz K, Furgang D, Fairlie K, Ferrandiz J, Nasri C, McKiernan M, Gunsolley J. Aggregatibacter actinomycetemcomitans and its relationship to initiation of localized aggressive periodontitis: longitudinal cohort study of initially healthy adolescents. J Clin Microbiol. 2007;45(12):3859–3869. - PMC - PubMed
-
- Zambon JJ, Christersson LA, Slots J. Actinobacillus actinomycetemcomitans in human periodontal disease. Prevalence in patient groups and distribution of biotypes and serotypes within families. J Periodontol. 1983;54(12):707–711. - PubMed
-
- van Winkelhoff AJ, Slots J. Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis in nonoral infections. Periodontol 2000. 1999;20:122–135. - PubMed
-
- Henderson B, Ward JM, Ready D. Aggregatibacter (Actinobacillus) actinomycetemcomitans: a triple A* periodontopathogen? Periodontol 2000. 2010;54(1):78–105. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
