

Reconsidering an active role for G-actin in cytoskeletal regulation
- PMID: 29321224
- PMCID: PMC5818056
- DOI: 10.1242/jcs.203760
Reconsidering an active role for G-actin in cytoskeletal regulation
Abstract
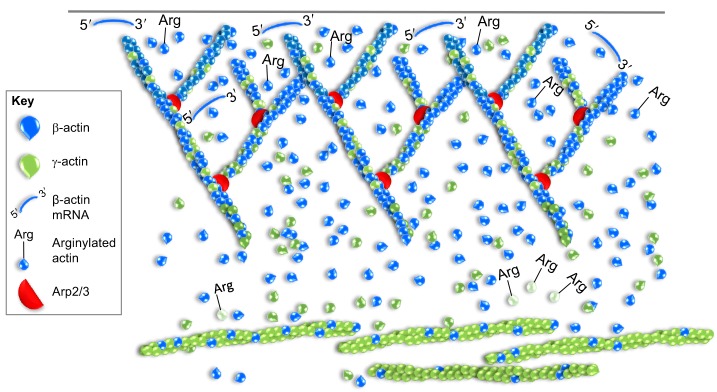
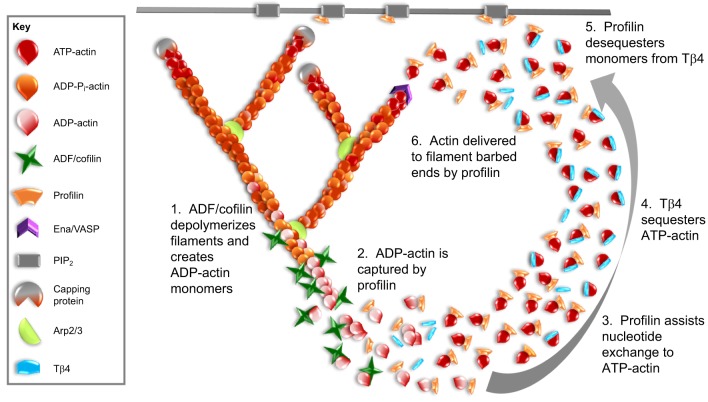
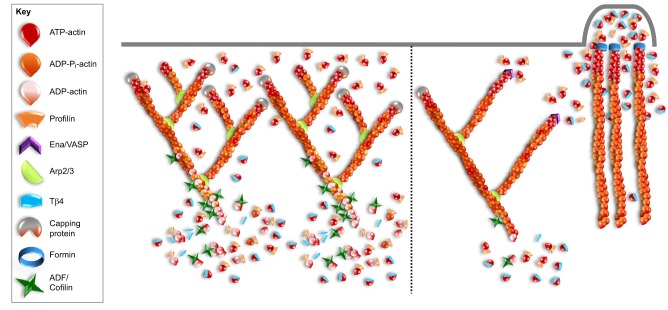
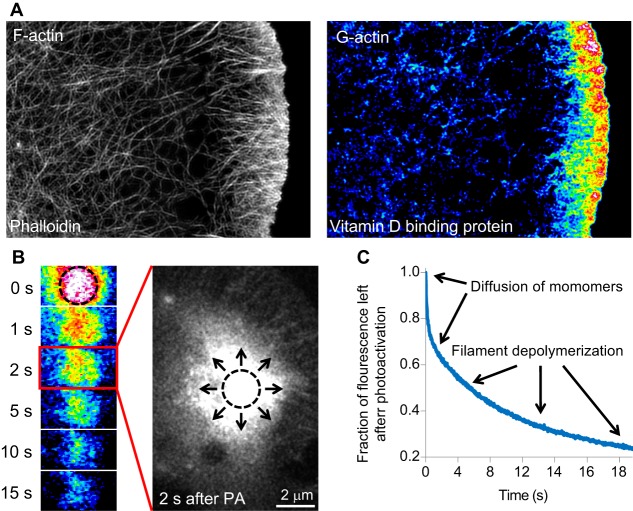
Globular (G)-actin, the actin monomer, assembles into polarized filaments that form networks that can provide structural support, generate force and organize the cell. Many of these structures are highly dynamic and to maintain them, the cell relies on a large reserve of monomers. Classically, the G-actin pool has been thought of as homogenous. However, recent work has shown that actin monomers can exist in distinct groups that can be targeted to specific networks, where they drive and modify filament assembly in ways that can have profound effects on cellular behavior. This Review focuses on the potential factors that could create functionally distinct pools of actin monomers in the cell, including differences between the actin isoforms and the regulation of G-actin by monomer binding proteins, such as profilin and thymosin β4. Owing to difficulties in studying and visualizing G-actin, our knowledge over the precise role that specific actin monomer pools play in regulating cellular actin dynamics remains incomplete. Here, we discuss some of these unanswered questions and also provide a summary of the methodologies currently available for the imaging of G-actin.
Keywords: G-actin; Profilin; Thymosin β 4; β-actin; γ-actin.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
