

Functional variants in the LRRK2 gene confer shared effects on risk for Crohn's disease and Parkinson's disease
- PMID: 29321258
- PMCID: PMC6028002
- DOI: 10.1126/scitranslmed.aai7795
Functional variants in the LRRK2 gene confer shared effects on risk for Crohn's disease and Parkinson's disease
Abstract
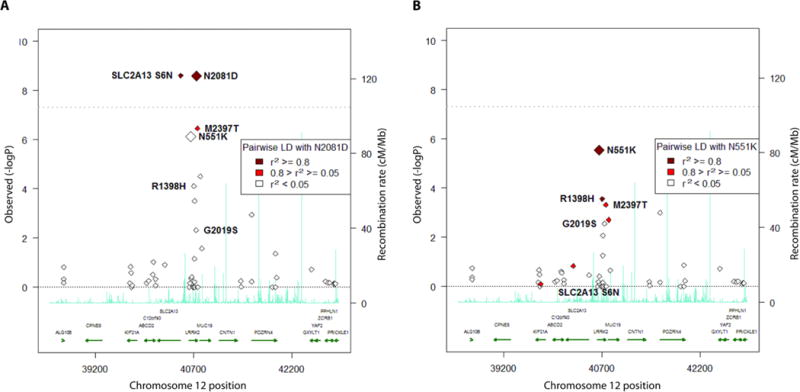
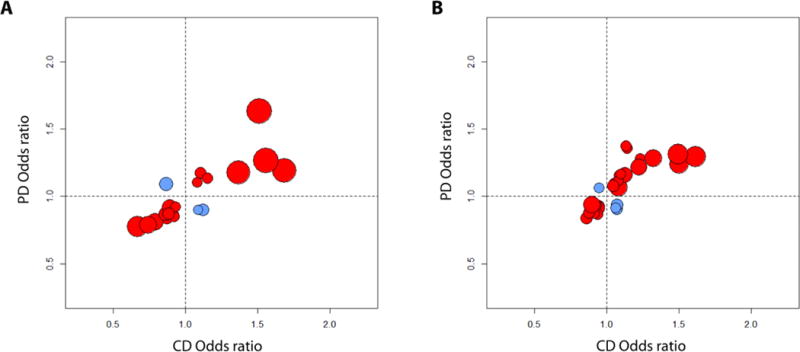
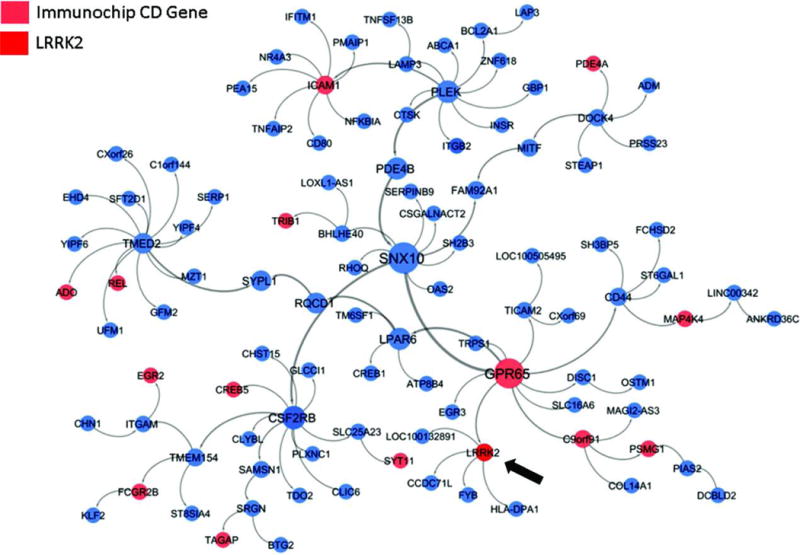
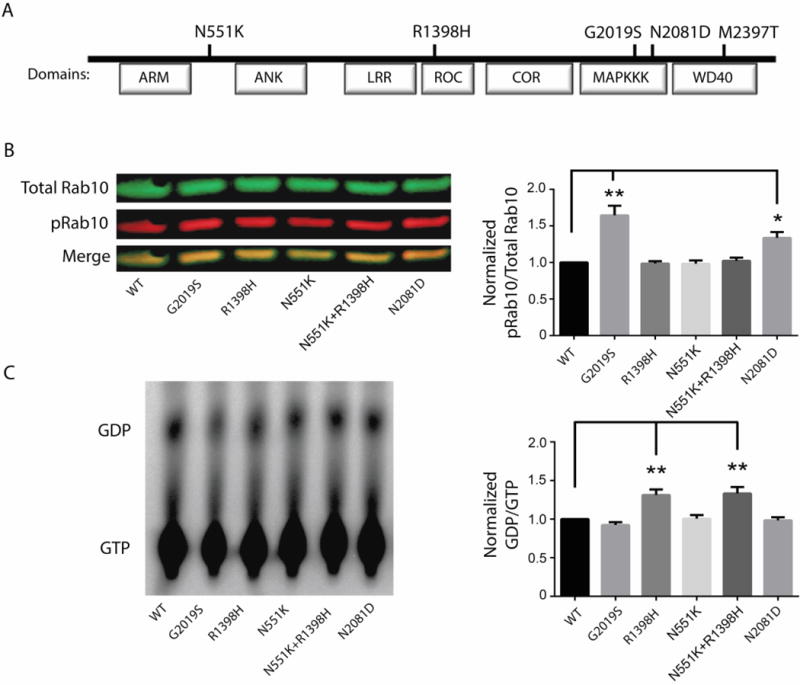
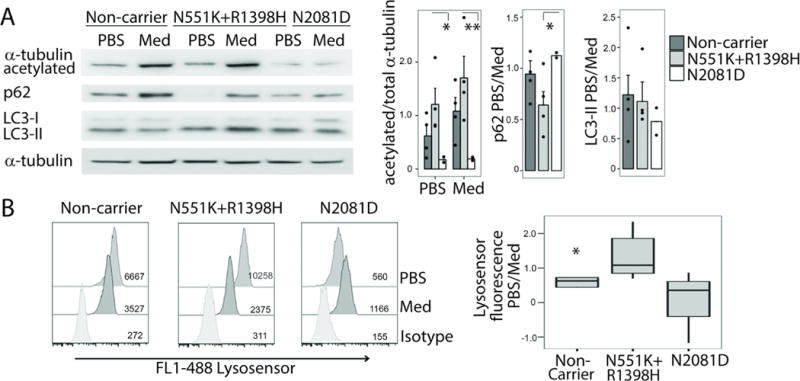
Crohn's disease (CD), a form of inflammatory bowel disease, has a higher prevalence in Ashkenazi Jewish than in non-Jewish European populations. To define the role of nonsynonymous mutations, we performed exome sequencing of Ashkenazi Jewish patients with CD, followed by array-based genotyping and association analysis in 2066 CD cases and 3633 healthy controls. We detected association signals in the LRRK2 gene that conferred risk for CD (N2081D variant, P = 9.5 × 10-10) or protection from CD (N551K variant, tagging R1398H-associated haplotype, P = 3.3 × 10-8). These variants affected CD age of onset, disease location, LRRK2 activity, and autophagy. Bayesian network analysis of CD patient intestinal tissue further implicated LRRK2 in CD pathogenesis. Analysis of the extended LRRK2 locus in 24,570 CD cases, patients with Parkinson's disease (PD), and healthy controls revealed extensive pleiotropy, with shared genetic effects between CD and PD in both Ashkenazi Jewish and non-Jewish cohorts. The LRRK2 N2081D CD risk allele is located in the same kinase domain as G2019S, a mutation that is the major genetic cause of familial and sporadic PD. Like the G2019S mutation, the N2081D variant was associated with increased kinase activity, whereas neither N551K nor R1398H variants on the protective haplotype altered kinase activity. We also confirmed that R1398H, but not N551K, increased guanosine triphosphate binding and hydrolyzing enzyme (GTPase) activity, thereby deactivating LRRK2. The presence of shared LRRK2 alleles in CD and PD provides refined insight into disease mechanisms and may have major implications for the treatment of these two seemingly unrelated diseases.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
RJD has consulted for Amicus Therapeutics, Alexion Pharmaceuticals, Genzyme-Sanofi, Kiniksa Pharmaceuticals,
Mitsubishi-Tanabe, Synageva Pharmaceuticals,
Recordati Rare Diseases,
Sangamo Therapeutics, and has received royalties from Shire. Shire: Royalties
YI has consulted for Neurotrope, Inc and Amathus Therapeutics, Inc.
SC has consulted for MyHeritage
ASC has consulted for AbbVie Pharmaceuticals, Janssen Pharmaceuticals, Takeda Pharmaceuticals, Pfeizer Pharmaceuticals, Ferring Pharmaceuticals, Miraca Life Sciences
DM has consulted for Janssen Pharmaceutical, UCB, Merck, Cidara, Qu Biologics.
The other authors declare no competing interests.
Data Availability: Samples from the Ashkenazi Genome Consortium are available from member institutions through a Material Transfer Agreement.
Figures
Comment in
-
Crohn's and Parkinson disease: is LRRK2 lurking around the corner?Nat Rev Gastroenterol Hepatol. 2018 Jun;15(6):330-331. doi: 10.1038/s41575-018-0006-9. Nat Rev Gastroenterol Hepatol. 2018. PMID: 29666431 No abstract available.
-
Brain and gut: Partners in crime.Mov Disord. 2018 Jul;33(7):1098. doi: 10.1002/mds.27407. Epub 2018 May 8. Mov Disord. 2018. PMID: 29737560 No abstract available.
References
-
- Jostins L, Ripke S, Weersma RK, Duerr RH, McGovern DP, Hui KY, Lee JC, Schumm LP, Sharma Y, Anderson CA, Essers J, Mitrovic M, Ning K, Cleynen I, Theatre E, Spain SL, Raychaudhuri S, Goyette P, Wei Z, Abraham C, Achkar JP, Ahmad T, Amininejad L, Ananthakrishnan AN, Andersen V, Andrews JM, Baidoo L, Balschun T, Bampton PA, Bitton A, Boucher G, Brand S, Buning C, Cohain A, Cichon S, D’Amato M, De Jong D, Devaney KL, Dubinsky M, Edwards C, Ellinghaus D, Ferguson LR, Franchimont D, Fransen K, Gearry R, Georges M, Gieger C, Glas J, Haritunians T, Hart A, Hawkey C, Hedl M, Hu X, Karlsen TH, Kupcinskas L, Kugathasan S, Latiano A, Laukens D, Lawrance IC, Lees CW, Louis E, Mahy G, Mansfield J, Morgan AR, Mowat C, Newman W, Palmieri O, Ponsioen CY, Potocnik U, Prescott NJ, Regueiro M, Rotter JI, Russell RK, Sanderson JD, Sans M, Satsangi J, Schreiber S, Simms LA, Sventoraityte J, Targan SR, Taylor KD, Tremelling M, Verspaget HW, De Vos M, Wijmenga C, Wilson DC, Winkelmann J, Xavier RJ, Zeissig S, Zhang B, Zhang CK, Zhao H, Silverberg MS, Annese V, Hakonarson H, Brant SR, Radford-Smith G, Mathew CG, Rioux JD, Schadt EE, Daly MJ, Franke A, Parkes M, Vermeire S, Barrett JC, Cho JH. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature. 2012;491:119–124. - PMC - PubMed
-
- Liu JZ, van Sommeren S, Huang H, Ng SC, Alberts R, Takahashi A, Ripke S, Lee JC, Jostins L, Shah T, Abedian S, Cheon JH, Cho J, Daryani NE, Franke L, Fuyuno Y, Hart A, Juyal RC, Juyal G, Kim WH, Morris AP, Poustchi H, Newman WG, Midha V, Orchard TR, Vahedi H, Sood A, Sung JJ, Malekzadeh R, Westra HJ, Yamazaki K, Yang SK, C. International Multiple Sclerosis Genetics, I. B. D. G. C. International. Barrett JC, Franke A, Alizadeh BZ, Parkes M, B KT, Daly MJ, Kubo M, Anderson CA, Weersma RK, I. B. D. G. Consortium Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat Genet. 2015;47:979–986. - PMC - PubMed
-
- Duerr RH, Taylor KD, Brant SR, Rioux JD, Silverberg MS, Daly MJ, Steinhart AH, Abraham C, Regueiro M, Griffiths A, Dassopoulos T, Bitton A, Yang H, Targan S, Datta LW, Kistner EO, Schumm LP, Lee AT, Gregersen PK, Barmada MM, Rotter JI, Nicolae DL, Cho JH. A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science. 2006;314:1461–1463. - PMC - PubMed
-
- Singh S, Kroe-Barrett RR, Canada KA, Zhu X, Sepulveda E, Wu H, He Y, Raymond EL, Ahlberg J, Frego LE, Amodeo LM, Catron KM, Presky DH, Hanke JH. Selective targeting of the IL23 pathway: Generation and characterization of a novel high-affinity humanized anti-IL23A antibody. mAbs. 2015;7:778–791. - PMC - PubMed
-
- Murthy A, Li Y, Peng I, Reichelt M, Katakam AK, Noubade R, Roose-Girma M, DeVoss J, Diehl L, Graham RR, van Lookeren Campagne M. A Crohn’s disease variant in Atg16l1 enhances its degradation by caspase 3. Nature. 2014;506:456–462. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P50 NS094733/NS/NINDS NIH HHS/United States
- R21 NS050487/NS/NINDS NIH HHS/United States
- R01 NS036960/NS/NINDS NIH HHS/United States
- T32 GM007205/GM/NIGMS NIH HHS/United States
- R01 DK106593/DK/NIDDK NIH HHS/United States
- U01 DK062423/DK/NIDDK NIH HHS/United States
- R01 NS036711/NS/NINDS NIH HHS/United States
- P50 NS071674/NS/NINDS NIH HHS/United States
- R01 DK092235/DK/NIDDK NIH HHS/United States
- R01 MH095458/MH/NIMH NIH HHS/United States
- R01 AG042188/AG/NIA NIH HHS/United States
- R01 CA141743/CA/NCI NIH HHS/United States
- U01 DK062413/DK/NIDDK NIH HHS/United States
- R56 NS037167/NS/NINDS NIH HHS/United States
- R01 MH084098/MH/NIMH NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- MR/L000261/1/MRC_/Medical Research Council/United Kingdom
- P01 AG021654/AG/NIA NIH HHS/United States
- U24 DK062429/DK/NIDDK NIH HHS/United States
- T32 HD007149/HD/NICHD NIH HHS/United States
- R01 HS021747/HS/AHRQ HHS/United States
- F30 DK098927/DK/NIDDK NIH HHS/United States
- U01 DK062429/DK/NIDDK NIH HHS/United States
- R01 NS060809/NS/NINDS NIH HHS/United States
- U01 DK062422/DK/NIDDK NIH HHS/United States
- U54 CA121852/CA/NCI NIH HHS/United States
- R01 NS060113/NS/NINDS NIH HHS/United States
- R01 NS037167/NS/NINDS NIH HHS/United States
- P30 AG038072/AG/NIA NIH HHS/United States
- RC2 MH089964/MH/NIMH NIH HHS/United States
- U01 AI067068/AI/NIAID NIH HHS/United States
- P01 DK046763/DK/NIDDK NIH HHS/United States
- R01 NS076843/NS/NINDS NIH HHS/United States
- U01 DK062420/DK/NIDDK NIH HHS/United States
- U01 DK062431/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
