

Gradual plasticity alters population dynamics in variable environments: thermal acclimation in the green alga Chlamydomonas reinhartdii
- PMID: 29321297
- PMCID: PMC5784192
- DOI: 10.1098/rspb.2017.1942
Gradual plasticity alters population dynamics in variable environments: thermal acclimation in the green alga Chlamydomonas reinhartdii
Abstract
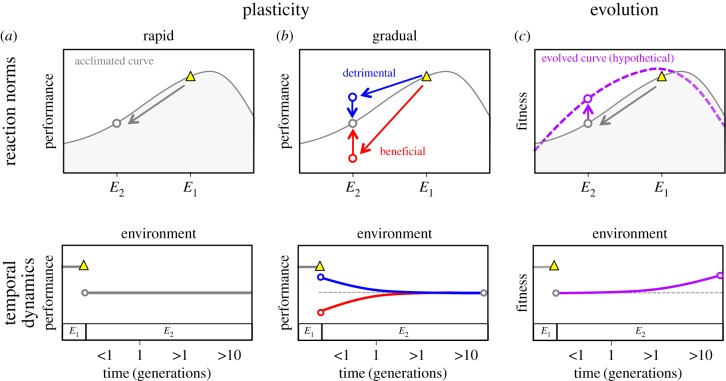
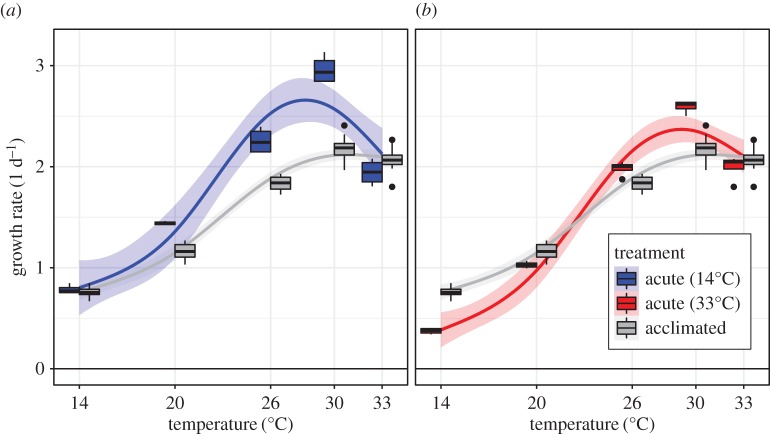
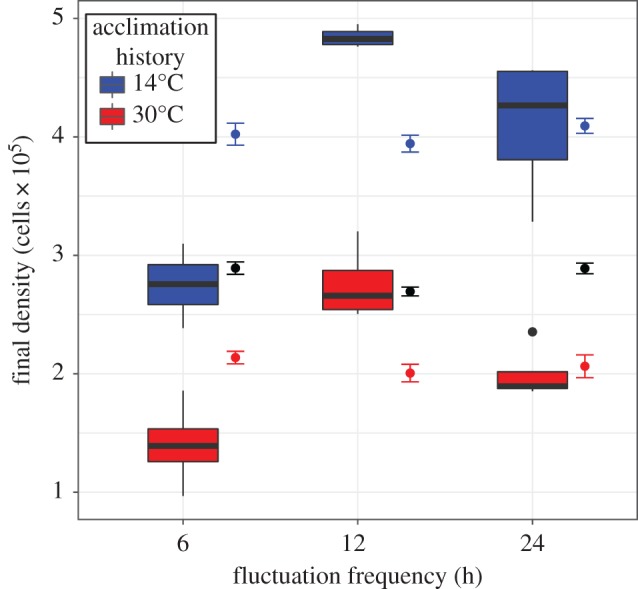
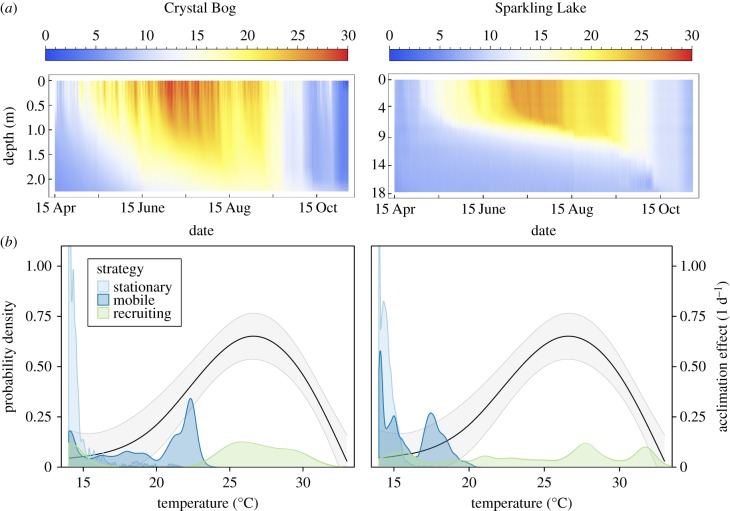
Environmental variability is ubiquitous, but its effects on populations are not fully understood or predictable. Recent attention has focused on how rapid evolution can impact ecological dynamics via adaptive trait change. However, the impact of trait change arising from plastic responses has received less attention, and is often assumed to optimize performance and unfold on a separate, faster timescale than ecological dynamics. Challenging these assumptions, we propose that gradual plasticity is important for ecological dynamics, and present a study of the plastic responses of the freshwater green algae Chlamydomonas reinhardtii as it acclimates to temperature changes. First, we show that C. reinhardtii's gradual acclimation responses can both enhance and suppress its performance after a perturbation, depending on its prior thermal history. Second, we demonstrate that where conventional approaches fail to predict the population dynamics of C. reinhardtii exposed to temperature fluctuations, a new model of gradual acclimation succeeds. Finally, using high-resolution data, we show that phytoplankton in lake ecosystems can experience thermal variation sufficient to make acclimation relevant. These results challenge prevailing assumptions about plasticity's interactions with ecological dynamics. Amidst the current emphasis on rapid evolution, it is critical that we also develop predictive methods accounting for plasticity.
Keywords: beneficial acclimation hypothesis; ecological forecasting; environmental variation; phenotypic plasticity; temperature; thermal performance.
© 2018 The Author(s).
Conflict of interest statement
We have no competing interests.
Figures

References
-
- Litchman E, Klausmeier CA. 2008. Trait-based community ecology of phytoplankton. Annu. Rev. Ecol. Evol. Syst. 39, 615–639. (10.1146/annurev.ecolsys.39.110707.173549) - DOI
-
- Kremer CT, Williams AK, Finiguerra M, Fong AA, Kellerman A, Paver SF, Tolar BB, Toscano BJ. 2017. Realizing the potential of trait-based aquatic ecology: new tools and collaborative approaches. Limnol. Oceanogr. 62, 253–271. (10.1002/lno.10392) - DOI
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
