

Herpes Simplex Virus 1 Mutant with Point Mutations in UL39 Is Impaired for Acute Viral Replication in Mice, Establishment of Latency, and Explant-Induced Reactivation
- PMID: 29321311
- PMCID: PMC5972887
- DOI: 10.1128/JVI.01654-17
Herpes Simplex Virus 1 Mutant with Point Mutations in UL39 Is Impaired for Acute Viral Replication in Mice, Establishment of Latency, and Explant-Induced Reactivation
Abstract
In the process of generating herpes simplex virus 1 (HSV-1) mutations in the viral regulatory gene encoding infected cell protein 0 (ICP0), we isolated a viral mutant, termed KOS-NA, that was severely impaired for acute replication in the eyes and trigeminal ganglia (TG) of mice, defective in establishing a latent infection, and reactivated poorly from explanted TG. To identify the secondary mutation(s) responsible for the impaired phenotypes of this mutant, we sequenced the KOS-NA genome and noted that it contained two nonsynonymous mutations in UL39, which encodes the large subunit of ribonucleotide reductase, ICP6. These mutations resulted in lysine-to-proline (residue 393) and arginine-to-histidine (residue 950) substitutions in ICP6. To determine whether alteration of these amino acids was responsible for the KOS-NA phenotypes in vivo, we recombined the wild-type UL39 gene into the KOS-NA genome and rescued its acute replication phenotypes in mice. To further establish the role of UL39 in KOS-NA's decreased pathogenicity, the UL39 mutations were recombined into HSV-1 (generating UL39mut), and this mutant virus showed reduced ocular and TG replication in mice comparable to that of KOS-NA. Interestingly, ICP6 protein levels were reduced in KOS-NA-infected cells relative to the wild-type protein. Moreover, we observed that KOS-NA does not counteract caspase 8-induced apoptosis, unlike wild-type strain KOS. Based on alignment studies with other HSV-1 ICP6 homologs, our data suggest that amino acid 950 of ICP6 likely plays an important role in ICP6 accumulation and inhibition of apoptosis, consequently impairing HSV-1 pathogenesis in a mouse model of HSV-1 infection.IMPORTANCE HSV-1 is a major human pathogen that infects ∼80% of the human population and can be life threatening to infected neonates or immunocompromised individuals. Effective therapies for treatment of recurrent HSV-1 infections are limited, which emphasizes a critical need to understand in greater detail the events that modulate HSV-1 replication and pathogenesis. In the current study, we identified a neuroattenuated HSV-1 mutant (i.e., KOS-NA) that contains novel mutations in the UL39 gene, which codes for the large subunit of ribonucleotide reductase (also known as ICP6). This mutant form of ICP6 was responsible for the attenuation of KOS-NA in vivo and resulted in diminished ICP6 protein levels and antiapoptotic effect. Thus, we have determined that subtle alteration of the UL39 gene regulates expression and functions of ICP6 and severely impacts HSV-1 pathogenesis, potentially making KOS-NA a promising vaccine candidate against HSV-1.
Keywords: HSV-1; ICP6; UL39; antiapoptosis; pathogenesis; ribonucleotide reductase; viral mutant.
Copyright © 2018 American Society for Microbiology.
Figures
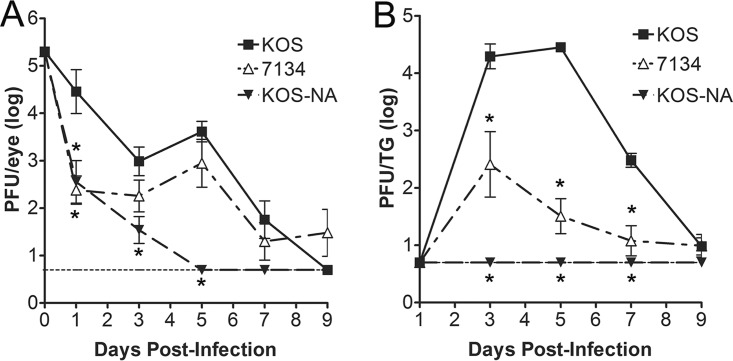
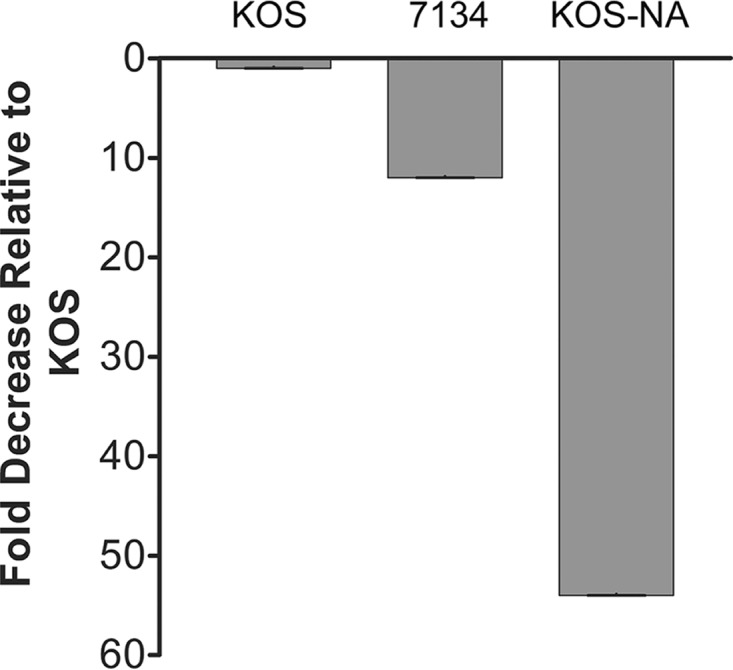





References
-
- Roizman B, Knipe DM, Whitley RJ. 2007. Herpes simplex viruses, p 2501–2601. In Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, vol 2 Lippincott Williams & Wilkins, Philadelphia, PA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
