

Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells
- PMID: 29321591
- PMCID: PMC5762922
- DOI: 10.1038/s41598-017-18672-5
Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells
Abstract
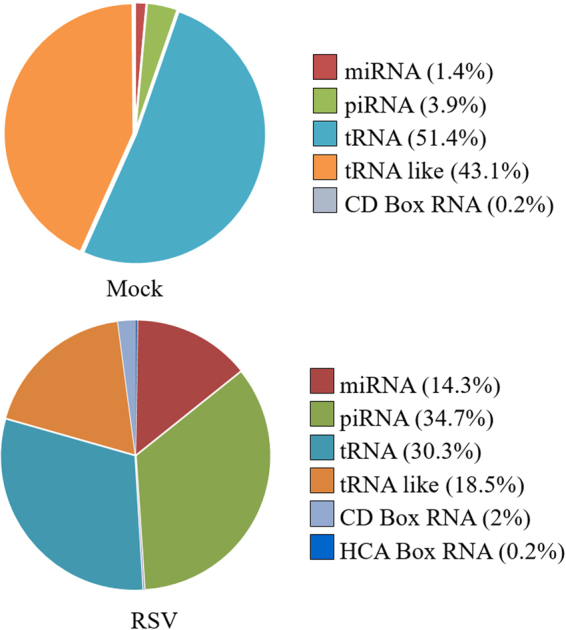
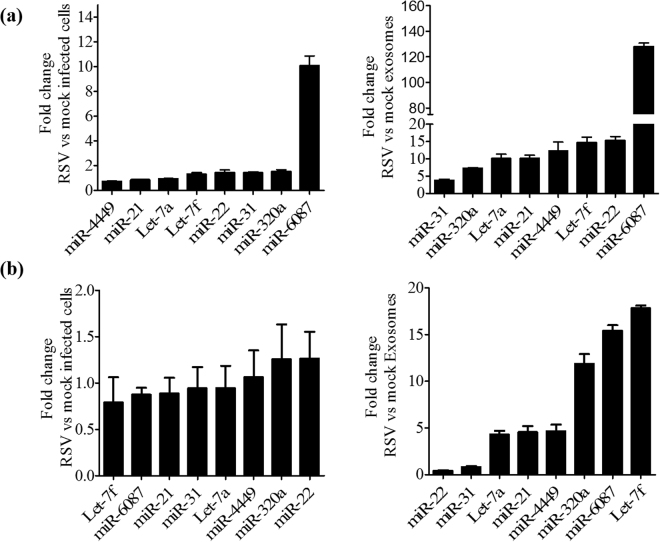
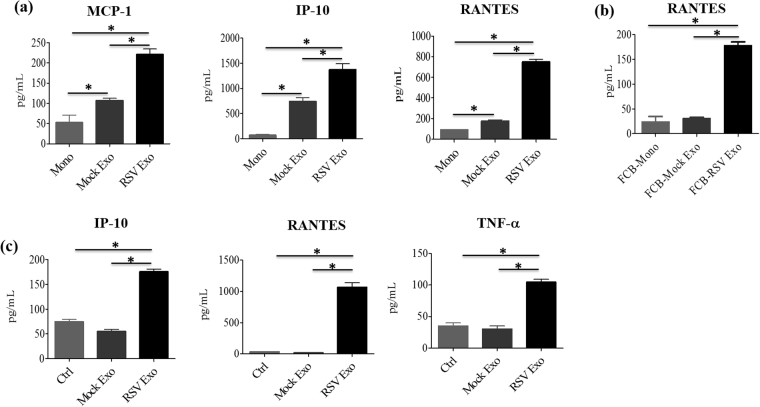
Exosomes are microvesicles known to carry biologically active molecules, including RNA, DNA and proteins. Viral infections can induce profound changes in exosome composition, and exosomes have been implicated in viral transmission and pathogenesis. No information is current available regarding exosome composition and function during infection with Respiratory Syncytial Virus (RSV), the most important cause of lower respiratory tract infections in children. In this study, we characterized exosomes released from RSV-infected lung carcinoma-derived A549 cells. RNA deep sequencing revealed that RSV exosomes contain a diverse range of RNA species like messenger and ribosomal RNA fragments, as well as small noncoding RNAs, in a proportion different from exosomes isolated from mock-infected cells. We observed that both RNA and protein signatures of RSV were present in exosomes, however, they were not able to establish productive infection in uninfected cells. Exosomes isolated from RSV-infected cells were able to activate innate immune response by inducing cytokine and chemokine release from human monocytes and airway epithelial cells. These data suggest that exosomes may play an important role in pathogenesis or protection against disease, therefore understating their role in RSV infection may open new avenues for target identification and development of novel therapeutics.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Harding C, Heuser J, Stahl P. Endocytosis and intracellular processing of transferrin and colloidal gold-transferrin in rat reticulocytes: demonstration of a pathway for receptor shedding. Eur. J. Cell Biol. 1984;35:256–263. - PubMed
-
- Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes) J. Biol. Chem. 1987;262:9412–9420. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
