

Fecal Microbiota Transplantation, Commensal Escherichia coli and Lactobacillus johnsonii Strains Differentially Restore Intestinal and Systemic Adaptive Immune Cell Populations Following Broad-spectrum Antibiotic Treatment
- PMID: 29321764
- PMCID: PMC5732213
- DOI: 10.3389/fmicb.2017.02430
Fecal Microbiota Transplantation, Commensal Escherichia coli and Lactobacillus johnsonii Strains Differentially Restore Intestinal and Systemic Adaptive Immune Cell Populations Following Broad-spectrum Antibiotic Treatment
Abstract
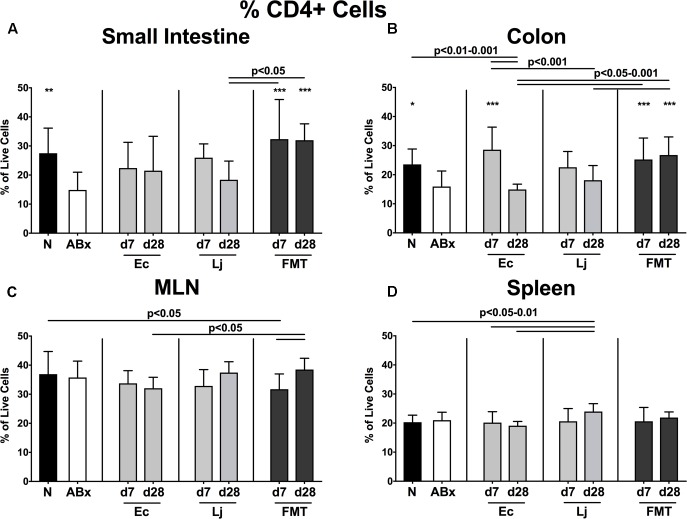
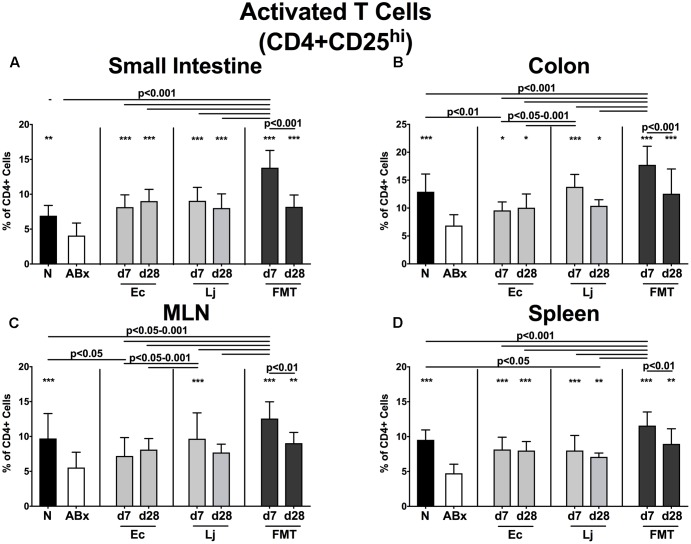
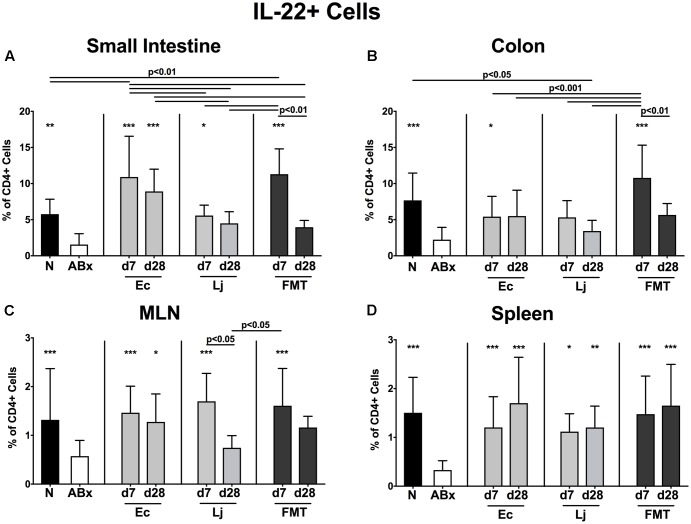
The essential role of the intestinal microbiota in the well-functioning of host immunity necessitates the investigation of species-specific impacts on this interplay. Aim of this study was to examine the ability of defined Gram-positive and Gram-negative intestinal commensal bacterial species, namely Escherichia coli and Lactobacillus johnsonii, respectively, to restore immune functions in mice that were immunosuppressed by antibiotics-induced microbiota depletion. Conventional mice were subjected to broad-spectrum antibiotic treatment for 8 weeks and perorally reassociated with E. coli, L. johnsonii or with a complex murine microbiota by fecal microbiota transplantation (FMT). Analyses at days (d) 7 and 28 revealed that immune cell populations in the small and large intestines, mesenteric lymph nodes and spleens of mice were decreased after antibiotic treatment but were completely or at least partially restored upon FMT or by recolonization with the respective bacterial species. Remarkably, L. johnsonii recolonization resulted in the highest CD4+ and CD8+ cell numbers in the small intestine and spleen, whereas neither of the commensal species could stably restore those cell populations in the colon until d28. Meanwhile less efficient than FMT, both species increased the frequencies of regulatory T cells and activated dendritic cells and completely restored intestinal memory/effector T cell populations at d28. Furthermore, recolonization with either single species maintained pro- and anti-inflammatory immune functions in parallel. However, FMT could most effectively recover the decreased frequencies of cytokine producing CD4+ lymphocytes in mucosal and systemic compartments. E. coli recolonization increased the production of cytokines such as TNF, IFN-γ, IL-17, and IL-22, particularly in the small intestine. Conversely, only L. johnsonii recolonization maintained colonic IL-10 production. In summary, FMT appears to be most efficient in the restoration of antibiotics-induced collateral damages to the immune system. However, defined intestinal commensals such as E. coli and L. johnsonii have the potential to restore individual functions of intestinal and systemic immunity. In conclusion, our data provide novel insights into the distinct role of individual commensal bacteria in maintaining immune functions during/following dysbiosis induced by antibiotic therapy thereby shaping host immunity and might thus open novel therapeutical avenues in conditions of perturbed microbiota composition.
Keywords: Escherichia coli; Lactobacillus johnsonii; commensal intestinal microbiota; fecal microbiota transplantation; immune restoration; immune-modulating effects; intestinal mucosal and peripheral and central immunity; secondary abiotic (gnotobiotic) mice.
Figures







References
-
- Bereswill S., Ekmekciu I., Escher U., Fiebiger U., Stingl K., Heimesaat M. M. (2017). Lactobacillus johnsonii ameliorates intestinal, extra-intestinal and systemic pro-inflammatory immune responses following murine Campylobacter jejuni infection. Sci. Rep. 7:2138. 10.1038/s41598-017-02436-2 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials