

Arabidopsis Basic Helix-Loop-Helix 34 (bHLH34) Is Involved in Glucose Signaling through Binding to a GAGA Cis-Element
- PMID: 29321786
- PMCID: PMC5732184
- DOI: 10.3389/fpls.2017.02100
Arabidopsis Basic Helix-Loop-Helix 34 (bHLH34) Is Involved in Glucose Signaling through Binding to a GAGA Cis-Element
Abstract
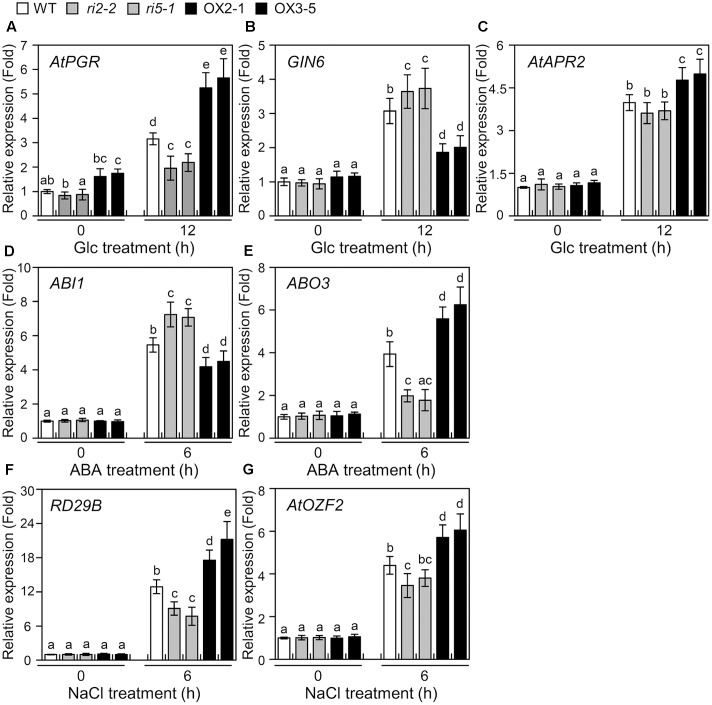
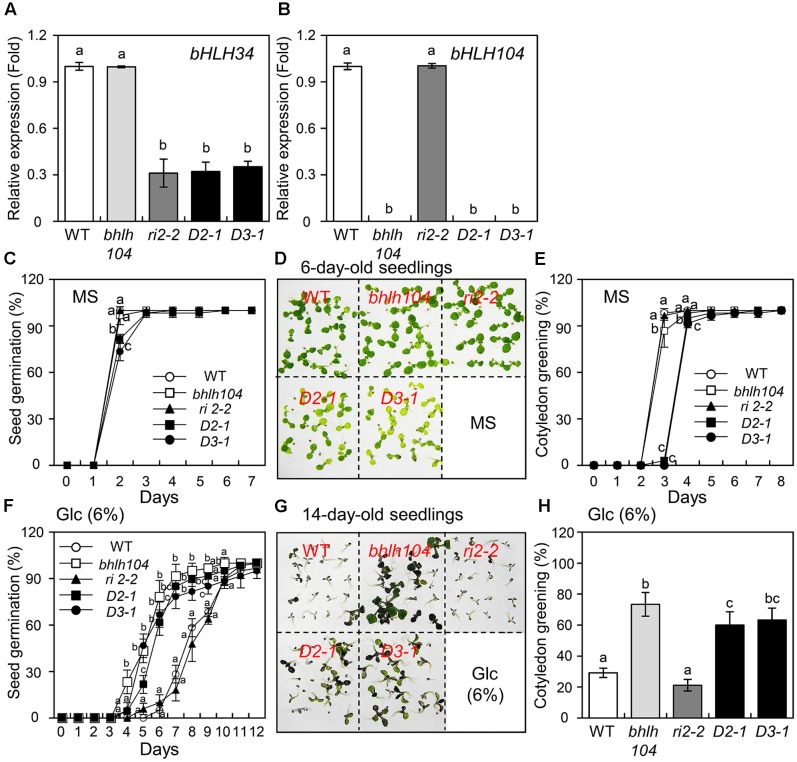
The modulation of glucose (Glc) homeostasis and signaling is crucial for plant growth and development. Nevertheless, the molecular signaling mechanism by which a plant senses a cellular Glc level and coordinates the expression of Glc-responsive genes is still incompletely understood. Previous studies have shown that Arabidopsis thaliana plasma membrane Glc-responsive regulator (AtPGR) is a component of the Glc-responsive pathway. Here, we demonstrated that a transcription factor bHLH34 binds to 5'-GAGA-3' element of the promoter region of AtPGR in vitro, and activates beta-glucuronidase (GUS) activity upon Glc treatment in AtPGR promoter-GUS transgenic plants. Gain- and loss-of-function analyses suggested that the bHLH34 involved in the responses to not only Glc, but also abscisic acid (ABA) and salinity. These results suggest that bHLH34 functions as a transcription factor in the Glc-mediated stress responsive pathway as well as an activator of AtPGR transcription. Furthermore, genetic experiments revealed that in Glc response, the functions of bHLH34 are different from that of a bHLH104, a homolog of bHLH34. Collectively, our findings indicate that bHLH34 is a positive regulator of Glc, and may affect ABA or salinity response, whereas bHLH104 is a negative regulator and epistatic to bHLH34 in the Glc response.
Keywords: AtPGR; activator; bHLH34; glucose-responsive element; transcription factor.
Figures




References
-
- Arenas-Huertero F., Arroyo A., Zhou L., Sheen J., León P. (2000). Analysis of Arabidopsis glucose insensitive mutants, gin5 and gin6, reveals a central role of the plant hormone ABA in the regulation of plant vegetative development by sugar. Genes Dev. 14 2085–2096. 10.1101/gad.14.16.2085 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases