

Integrating restriction site-associated DNA sequencing (RAD-seq) with morphological cladistic analysis clarifies evolutionary relationships among major species groups of bee orchids
- PMID: 29325077
- PMCID: PMC5786241
- DOI: 10.1093/aob/mcx129
Integrating restriction site-associated DNA sequencing (RAD-seq) with morphological cladistic analysis clarifies evolutionary relationships among major species groups of bee orchids
Abstract
Background and aims: Bee orchids (Ophrys) have become the most popular model system for studying reproduction via insect-mediated pseudo-copulation and for exploring the consequent, putatively adaptive, evolutionary radiations. However, despite intensive past research, both the phylogenetic structure and species diversity within the genus remain highly contentious. Here, we integrate next-generation sequencing and morphological cladistic techniques to clarify the phylogeny of the genus.
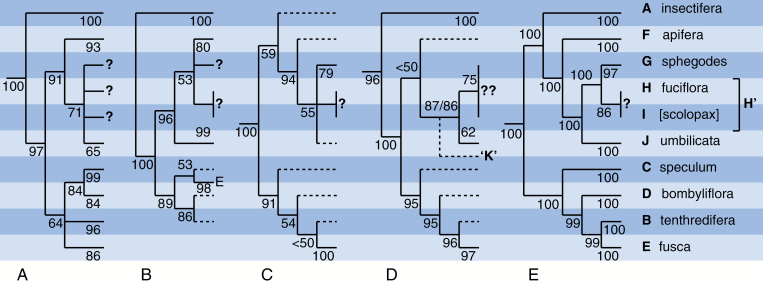
Methods: At least two accessions of each of the ten species groups previously circumscribed from large-scale cloned nuclear ribosomal internal transcibed spacer (nrITS) sequencing were subjected to restriction site-associated sequencing (RAD-seq). The resulting matrix of 4159 single nucleotide polymorphisms (SNPs) for 34 accessions was used to construct an unrooted network and a rooted maximum likelihood phylogeny. A parallel morphological cladistic matrix of 43 characters generated both polymorphic and non-polymorphic sets of parsimony trees before being mapped across the RAD-seq topology.
Key results: RAD-seq data strongly support the monophyly of nine out of ten groups previously circumscribed using nrITS and resolve three major clades; in contrast, supposed microspecies are barely distinguishable. Strong incongruence separated the RAD-seq trees from both the morphological trees and traditional classifications; mapping of the morphological characters across the RAD-seq topology rendered them far more homoplastic.
Conclusions: The comparatively high level of morphological homoplasy reflects extensive convergence, whereas the derived placement of the fusca group is attributed to paedomorphic simplification. The phenotype of the most recent common ancestor of the extant lineages is inferred, but it post-dates the majority of the character-state changes that typify the genus. RAD-seq may represent the high-water mark of the contribution of molecular phylogenetics to understanding evolution within Ophrys; further progress will require large-scale population-level studies that integrate phenotypic and genotypic data in a cogent conceptual framework.
Keywords: Biogeography; Mediterranean; Ophrys; RAD-seq; character mapping; coalescence; convergence; evolution; internal transcribed spacer; macrospecies; microspecies; morphology; paedomorphosis; phylogenetics; plastid; pseudo-copulation; systematics.
© The Author(s) 2018. Published by Oxford University Press on behalf of the Annals of Botany Company.
Figures





References
-
- Alberch P, Gould SJ, Oster GF, Wake DB. 1979. Size and shape in ontogeny and phylogeny. Paleobiology 5: 269–317.
-
- Ayasse M, Gögler J, Stökl J. 2010. Pollinator-driven speciation in sexually deceptive orchids of the genus Ophrys. In: Glaubrecht M, ed. Evolution in action. Berlin: Springer, 101–118.
-
- Ayasse M, Schiestl FP, Paulus HF et al. . 2000. Evolution of reproductive strategies in the sexually deceptive orchid Ophrys sphegodes: how does flower-specific variation of odor signals influence reproductive success?Evolution 54: 1999–2006. - PubMed
-
- Bateman RM. 1994. Evolutionary–developmental change in the growth architecture of fossil rhizomorphic lycopsids: scenarios constructed on cladistic foundations. Biological Reviews 69: 527–597.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
