

The Mitochondrial GTPase Gem1 Contributes to the Cell Wall Stress Response and Invasive Growth of Candida albicans
- PMID: 29326680
- PMCID: PMC5742345
- DOI: 10.3389/fmicb.2017.02555
The Mitochondrial GTPase Gem1 Contributes to the Cell Wall Stress Response and Invasive Growth of Candida albicans
Abstract
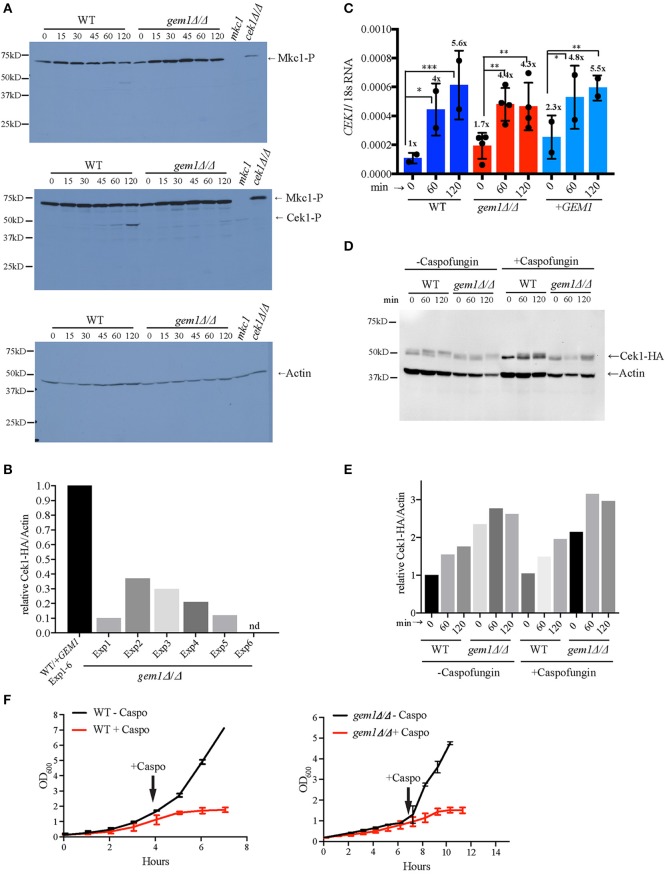
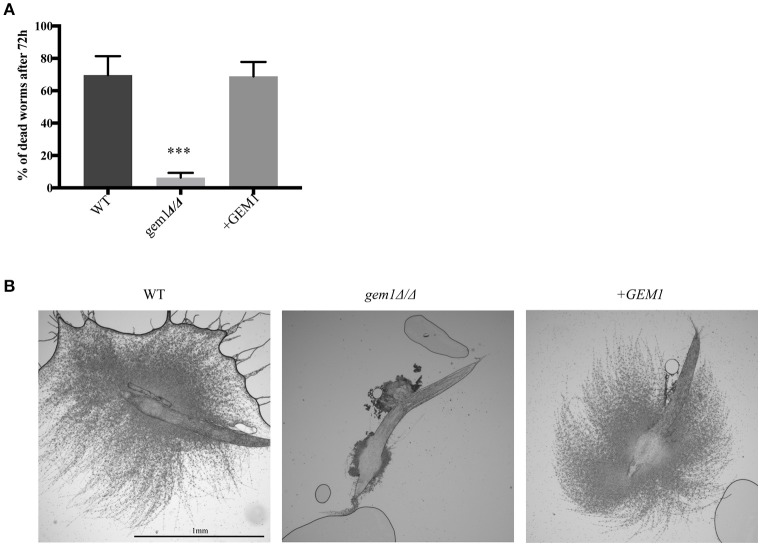
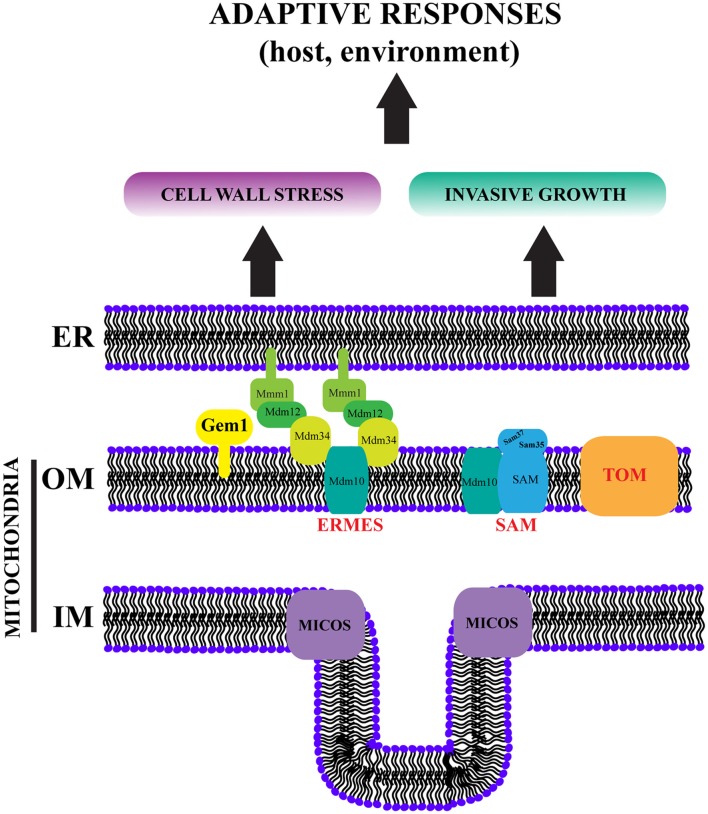
The interactions of mitochondria with the endoplasmic reticulum (ER) are crucial for maintaining proper mitochondrial morphology, function and dynamics. This enables cells to utilize their mitochondria optimally for energy production and anabolism, and it further provides for metabolic control over developmental decisions. In fungi, a key mechanism by which ER and mitochondria interact is via a membrane tether, the protein complex ERMES (ER-Mitochondria Encounter Structure). In the model yeast Saccharomyces cerevisiae, the mitochondrial GTPase Gem1 interacts with ERMES, and it has been proposed to regulate its activity. Here we report on the first characterization of Gem1 in a human fungal pathogen. We show that in Candida albicans Gem1 has a dominant role in ensuring proper mitochondrial morphology, and our data is consistent with Gem1 working with ERMES in this role. Mitochondrial respiration and steady state cellular phospholipid homeostasis are not impacted by inactivation of GEM1 in C. albicans. There are two major virulence-related consequences of disrupting mitochondrial morphology by GEM1 inactivation: C. albicans becomes hypersusceptible to cell wall stress, and is unable to grow invasively. In the gem1Δ/Δ mutant, it is specifically the invasive capacity of hyphae that is compromised, not the ability to transition from yeast to hyphal morphology, and this phenotype is shared with ERMES mutants. As a consequence of the hyphal invasion defect, the gem1Δ/Δ mutant is drastically hypovirulent in the worm infection model. Activation of the mitogen activated protein (MAP) kinase Cek1 is reduced in the gem1Δ/Δ mutant, and this function could explain both the susceptibility to cell wall stress and lack of invasive growth. This result establishes a new, respiration-independent mechanism of mitochondrial control over stress signaling and hyphal functions in C. albicans. We propose that ER-mitochondria interactions and the ER-Mitochondria Organizing Network (ERMIONE) play important roles in adaptive responses in fungi, in particular cell surface-related mechanisms that drive invasive growth and stress responsive behaviors that support fungal pathogenicity.
Keywords: CEK1; Candida albicans; invasive growth; mitochondria; virulence.
Figures




References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials