

Transmembrane Pickets Connect Cyto- and Pericellular Skeletons Forming Barriers to Receptor Engagement
- PMID: 29328918
- PMCID: PMC5929997
- DOI: 10.1016/j.cell.2017.12.023
Transmembrane Pickets Connect Cyto- and Pericellular Skeletons Forming Barriers to Receptor Engagement
Abstract
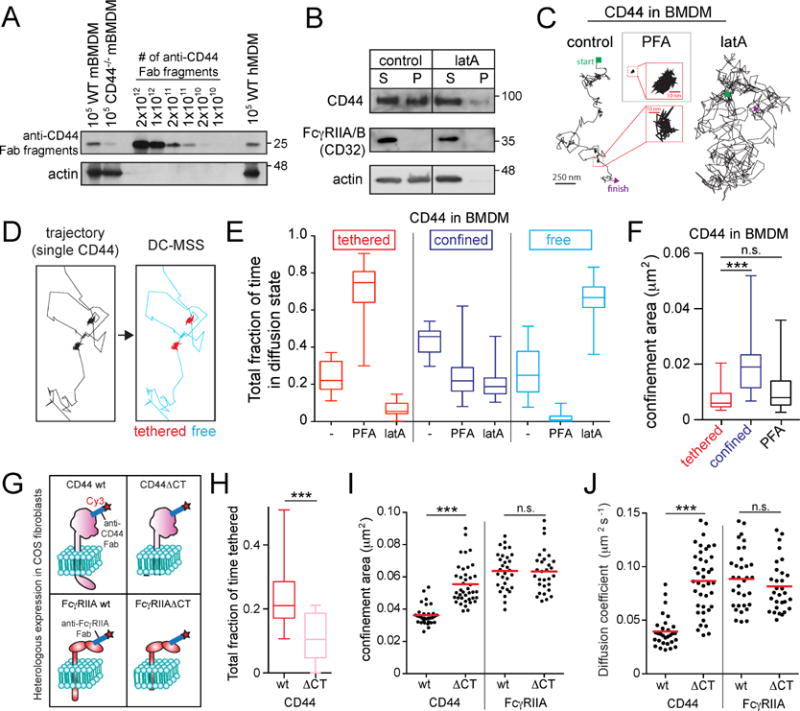
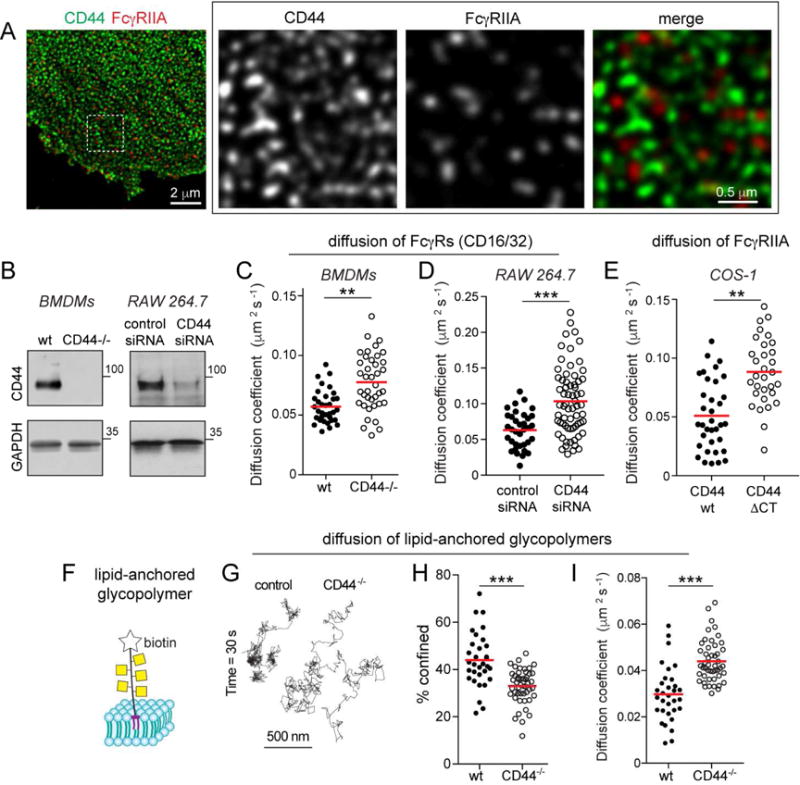
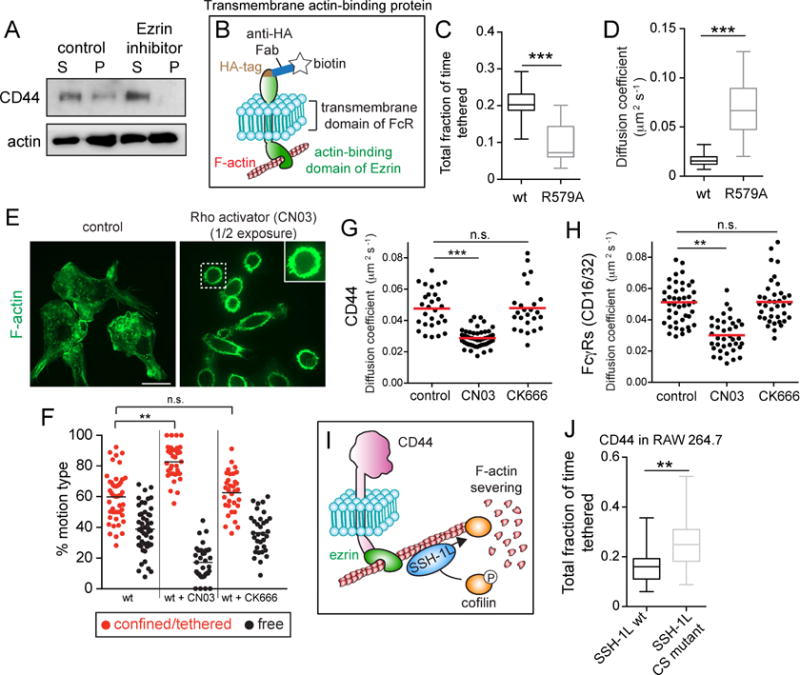
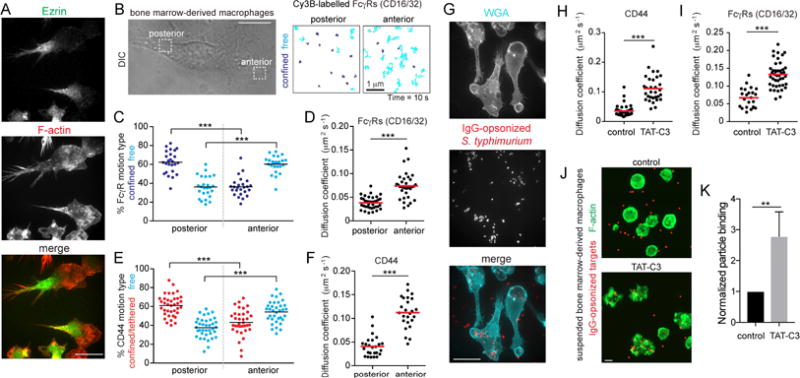
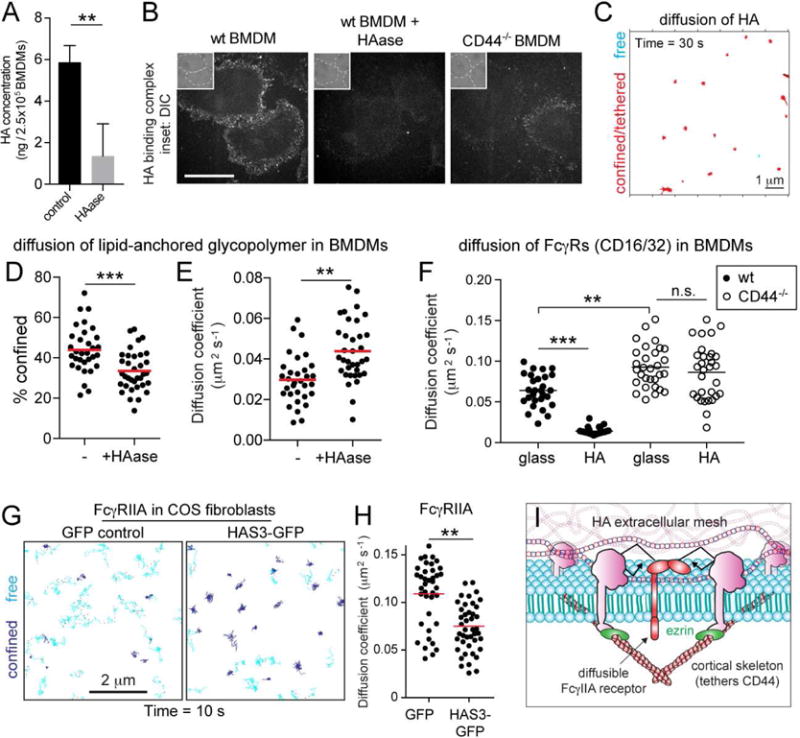
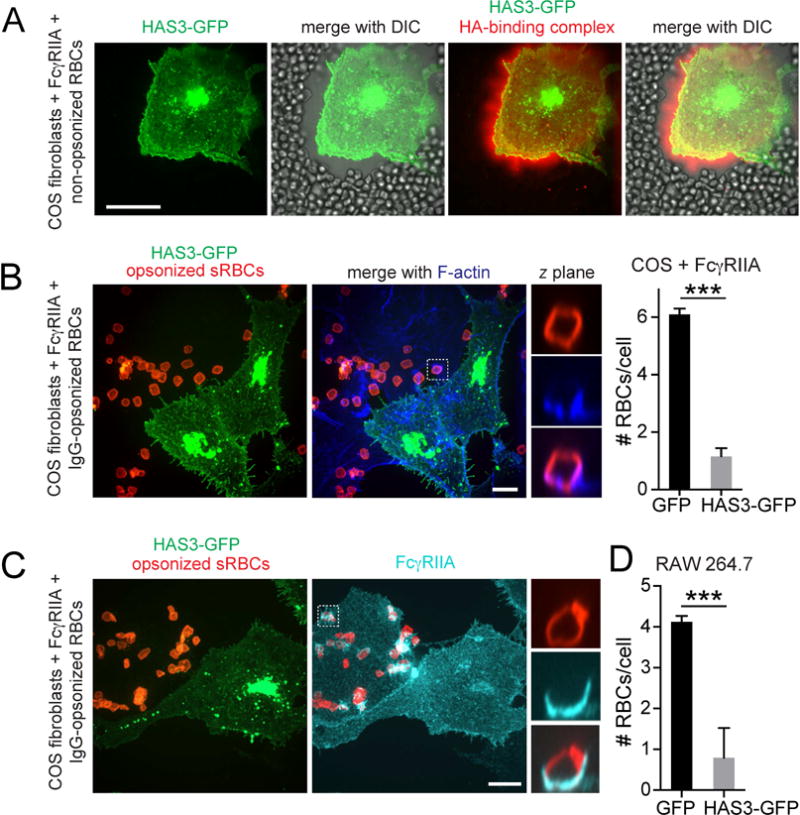
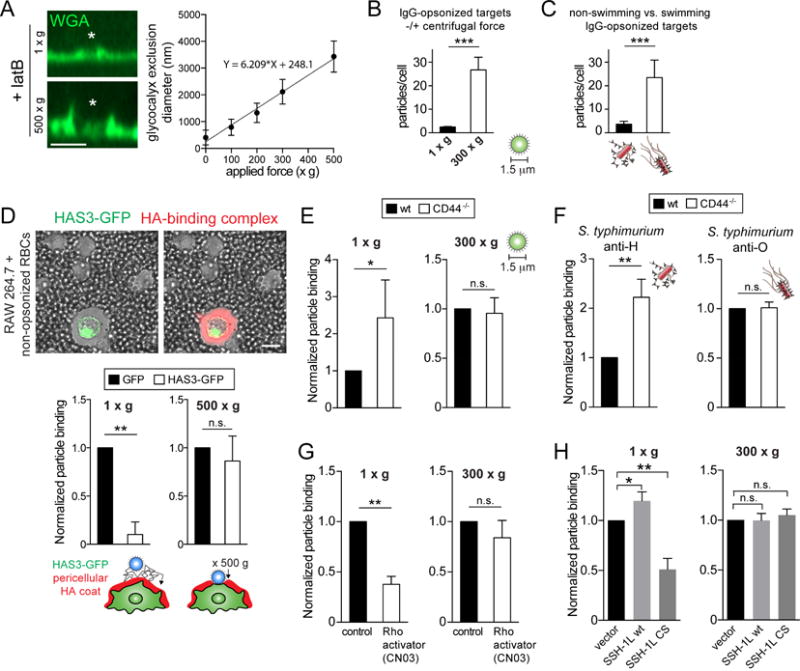
Phagocytic receptors must diffuse laterally to become activated upon clustering by multivalent targets. Receptor diffusion, however, can be obstructed by transmembrane proteins ("pickets") that are immobilized by interacting with the cortical cytoskeleton. The molecular identity of these pickets and their role in phagocytosis have not been defined. We used single-molecule tracking to study the interaction between Fcγ receptors and CD44, an abundant transmembrane protein capable of indirect association with F-actin, hence likely to serve as a picket. CD44 tethers reversibly to formin-induced actin filaments, curtailing receptor diffusion. Such linear filaments predominate in the trailing end of polarized macrophages, where receptor mobility was minimal. Conversely, receptors were most mobile at the leading edge, where Arp2/3-driven actin branching predominates. CD44 binds hyaluronan, anchoring a pericellular coat that also limits receptor displacement and obstructs access to phagocytic targets. Force must be applied to traverse the pericellular barrier, enabling receptors to engage their targets.
Keywords: Arp2/3; CD44; Fc receptor; diffusion barrier; ezrin; formin; glycocalyx; hyaluronan; phagocytosis; single particle tracking.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Aruffo A, Stamenkovic I, Melnick M, Underhill CB, Seed B. CD44 is the principal cell surface receptor for hyaluronate. Cell. 1990;61:1303–1313. - PubMed
-
- Downey GP, Botelho RJ, Butler JR, Moltyaner Y, Chien P, Schreiber AD, Grinstein S. Phagosomal maturation, acidification, and inhibition of bacterial growth in nonphagocytic cells transfected with FcgammaRIIA receptors. The Journal of biological chemistry. 1999;274:28436–28444. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
