

Preferential cholinergic excitation of corticopontine neurons
- PMID: 29330867
- PMCID: PMC5924837
- DOI: 10.1113/JP275194
Preferential cholinergic excitation of corticopontine neurons
Abstract
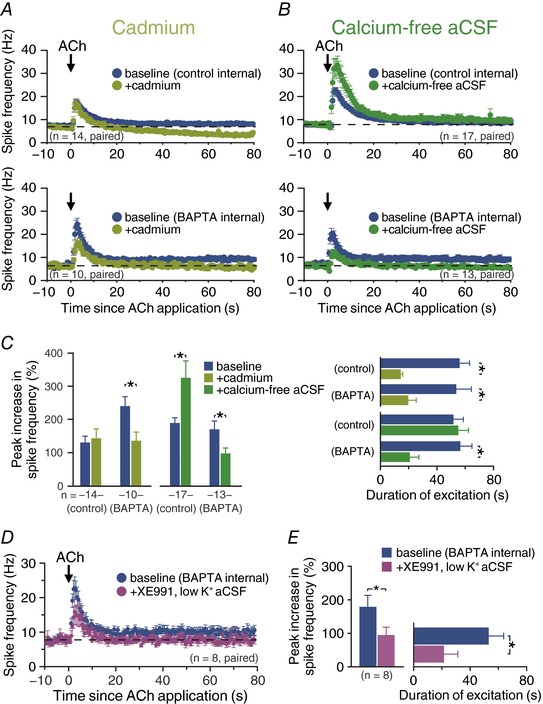
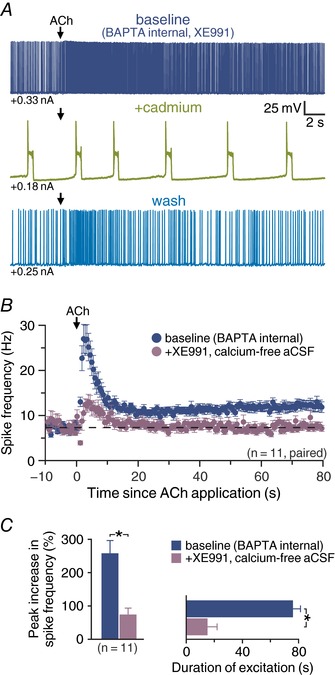
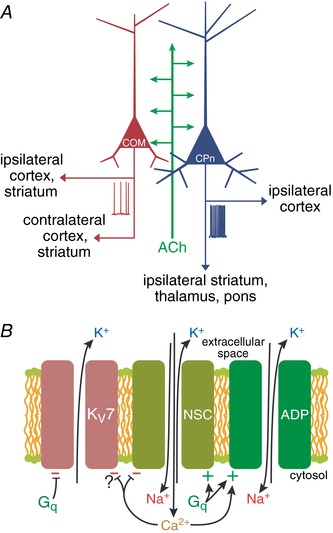
Key points: Phasic activation of M1 muscarinic receptors generates transient inhibition followed by longer lasting excitation in neocortical pyramidal neurons. Corticopontine neurons in the mouse prefrontal cortex exhibit weaker cholinergic inhibition, but more robust and longer lasting excitation, than neighbouring callosal projection neurons. Optogenetic release of endogenous ACh in response to single flashes of light (5 ms) preferentially enhances the excitability of corticopontine neurons for many tens of seconds. Cholinergic excitation of corticopontine neurons involves at least three ionic mechanisms: suppression of KV 7 currents, activation of the calcium-dependent non-specific cation conductance underlying afterdepolarizations, and activation of what appears to be a calcium-sensitive but calcium-permeable non-specific cation conductance. Preferential cholinergic excitation of prefrontal corticopontine neurons may facilitate top-down attentional processes and behaviours.
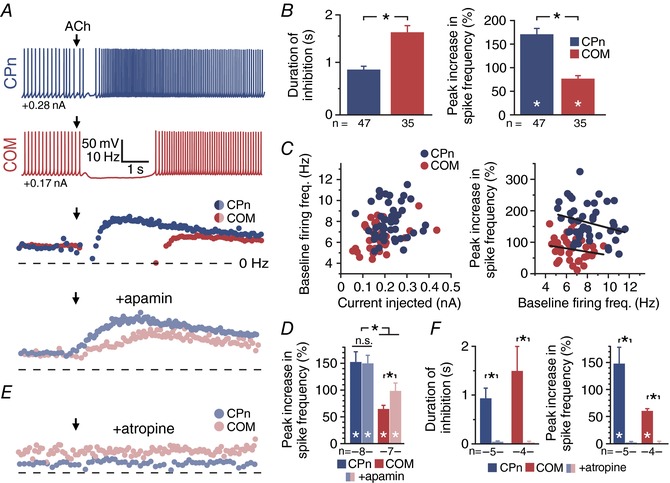
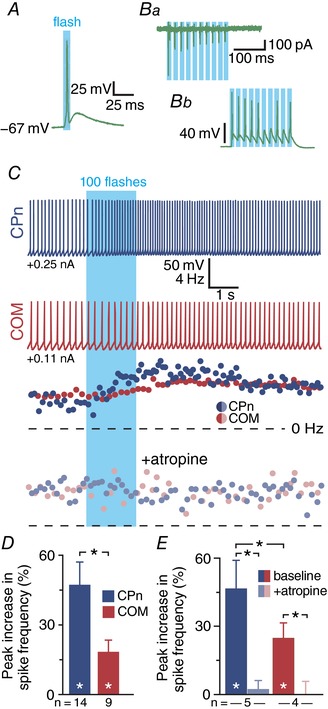
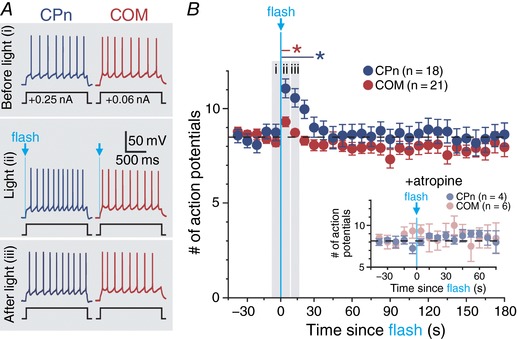
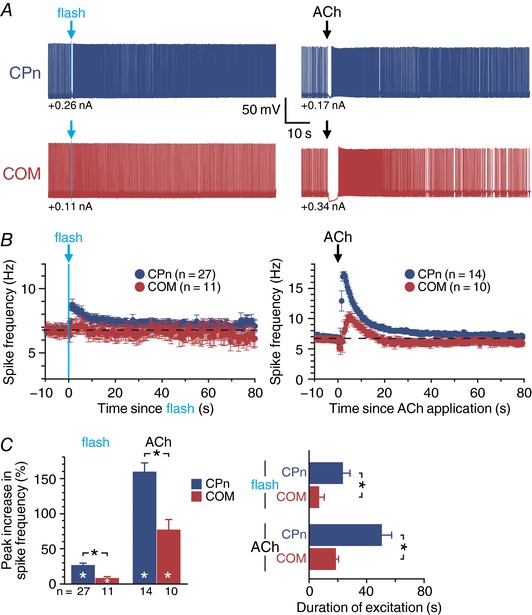
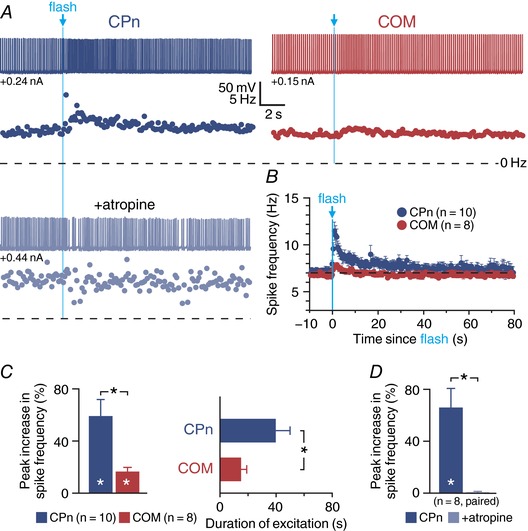
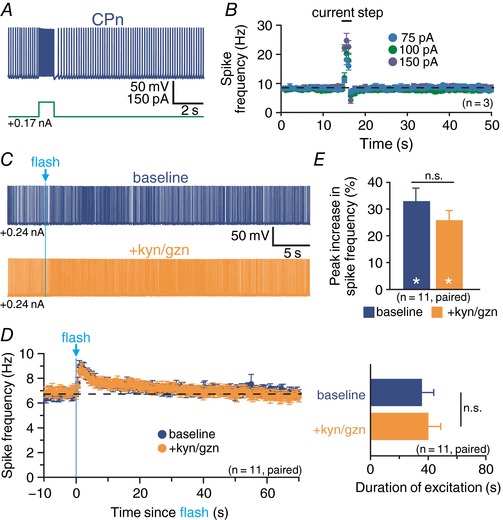
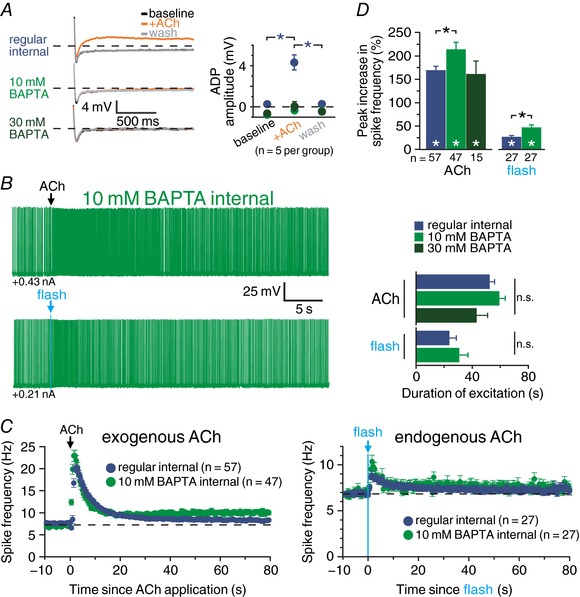
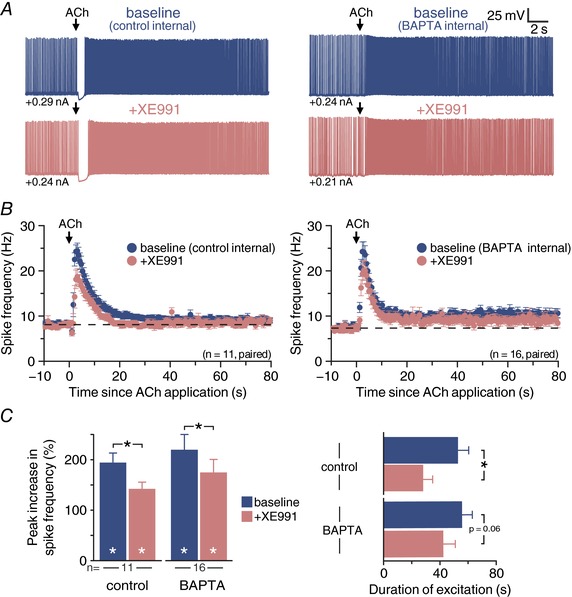
Abstract: Pyramidal neurons in layer 5 of the neocortex comprise two broad classes of projection neurons: corticofugal neurons, including corticopontine (CPn) neurons, and intratelencephalic neurons, including commissural/callosal (COM) neurons. These non-overlapping neuron subpopulations represent discrete cortical output channels contributing to perception, decision making and behaviour. CPn and COM neurons have distinct morphological and physiological characteristics, and divergent responses to modulatory transmitters such as serotonin and acetylcholine (ACh). To better understand how ACh regulates cortical output, in slices of mouse prefrontal cortex (PFC) we compared the responsivity of CPn and COM neurons to transient exposure to exogenous or endogenous ACh. In both neuron subtypes, exogenous ACh generated qualitatively similar biphasic responses in which brief hyperpolarization was followed by longer lasting enhancement of excitability. However, cholinergic inhibition was more pronounced in COM neurons, while excitatory responses were larger and longer lasting in CPn neurons. Similarly, optically triggered release of endogenous ACh from cholinergic terminals preferentially and persistently (for ∼40 s) enhanced the excitability of CPn neurons, but had little impact on COM neurons. Cholinergic excitation of CPn neurons involved at least three distinct ionic mechanisms: suppression of KV 7 channels (the 'M-current'), activation of the calcium-dependent non-specific cation conductance underlying afterdepolarizations, and activation of what appears to be a calcium-sensitive but calcium-permeable non-specific cation conductance. Our findings demonstrate projection-specific selectivity in cholinergic signalling in the PFC, and suggest that transient release of ACh during behaviour will preferentially promote corticofugal output.
Keywords: acetylcholine; neocortex; pyramidal neuron.
© 2018 The Authors. The Journal of Physiology © 2018 The Physiological Society.
Figures
Comment in
-
Ready, set, go: the bridging of attention to action by acetylcholine in prefrontal cortex.J Physiol. 2018 May 1;596(9):1539-1540. doi: 10.1113/JP275808. Epub 2018 Mar 26. J Physiol. 2018. PMID: 29512158 Free PMC article. No abstract available.
References
-
- Andrade R (1991). Cell excitation enhances muscarinic cholinergic responses in rat association cortex. Brain Res 548, 81–93. - PubMed
-
- Bentley P, Husain M & Dolan RJ (2004). Effects of cholinergic enhancement on visual stimulation, spatial attention, and spatial working memory. Neuron 41, 969–982. - PubMed
-
- Bockaert J, Perroy J, Becamel C, Marin P & Fagni L (2010). GPCR interacting proteins (GIPs) in the nervous system: Roles in physiology and pathologies. Annu Rev Pharmacol Toxicol 50, 89–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous