

Adaptive Immunity to Cryptococcus neoformans Infections
- PMID: 29333430
- PMCID: PMC5753166
- DOI: 10.3390/jof3040064
Adaptive Immunity to Cryptococcus neoformans Infections
Abstract
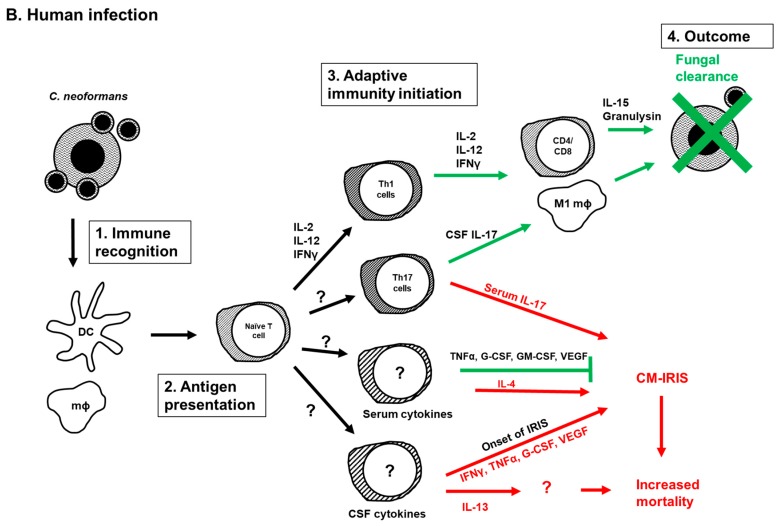
The Cryptococcus neoformans/Cryptococcus gattii species complex is a group of fungal pathogens with different phenotypic and genotypic diversity that cause disease in immunocompromised patients as well as in healthy individuals. The immune response resulting from the interaction between Cryptococcus and the host immune system is a key determinant of the disease outcome. The species C. neoformans causes the majority of human infections, and therefore almost all immunological studies focused on C. neoformans infections. Thus, this review presents current understanding on the role of adaptive immunity during C. neoformans infections both in humans and in animal models of disease.
Keywords: CD4; Cryptococcus; adaptive immunity; cytokines; dendritic cells; helper T cell type 1 (Th1); helper T cell type 17 (Th17); helper T cell type 2 (Th2); immune reconstitution inflammatory syndrome (IRIS).
Conflict of interest statement
Conflicts of Interest: The authors declare no conflict of interest.
Figures

References
-
- Pappas P.G., Perfect J.R., Cloud G.A., Larsen R.A., Pankey G.A., Lancaster D.J., Henderson H., Kauffman C.A., Haas D.W., Saccente M., et al. Cryptococcosis in human immunodeficiency virus-negative patients in the era of effective azole therapy. Clin. Infect. Dis. 2001;33:690–699. doi: 10.1086/322597. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
