

Unique metabolic activation of adipose tissue macrophages in obesity promotes inflammatory responses
- PMID: 29333574
- PMCID: PMC6448980
- DOI: 10.1007/s00125-017-4526-6
Unique metabolic activation of adipose tissue macrophages in obesity promotes inflammatory responses
Abstract
Aims/hypothesis: Recent studies have identified intracellular metabolism as a fundamental determinant of macrophage function. In obesity, proinflammatory macrophages accumulate in adipose tissue and trigger chronic low-grade inflammation, that promotes the development of systemic insulin resistance, yet changes in their intracellular energy metabolism are currently unknown. We therefore set out to study metabolic signatures of adipose tissue macrophages (ATMs) in lean and obese conditions.
Methods: F4/80-positive ATMs were isolated from obese vs lean mice. High-fat feeding of wild-type mice and myeloid-specific Hif1α-/- mice was used to examine the role of hypoxia-inducible factor-1α (HIF-1α) in ATMs part of obese adipose tissue. In vitro, bone marrow-derived macrophages were co-cultured with adipose tissue explants to examine adipose tissue-induced changes in macrophage phenotypes. Transcriptome analysis, real-time flux measurements, ELISA and several other approaches were used to determine the metabolic signatures and inflammatory status of macrophages. In addition, various metabolic routes were inhibited to determine their relevance for cytokine production.
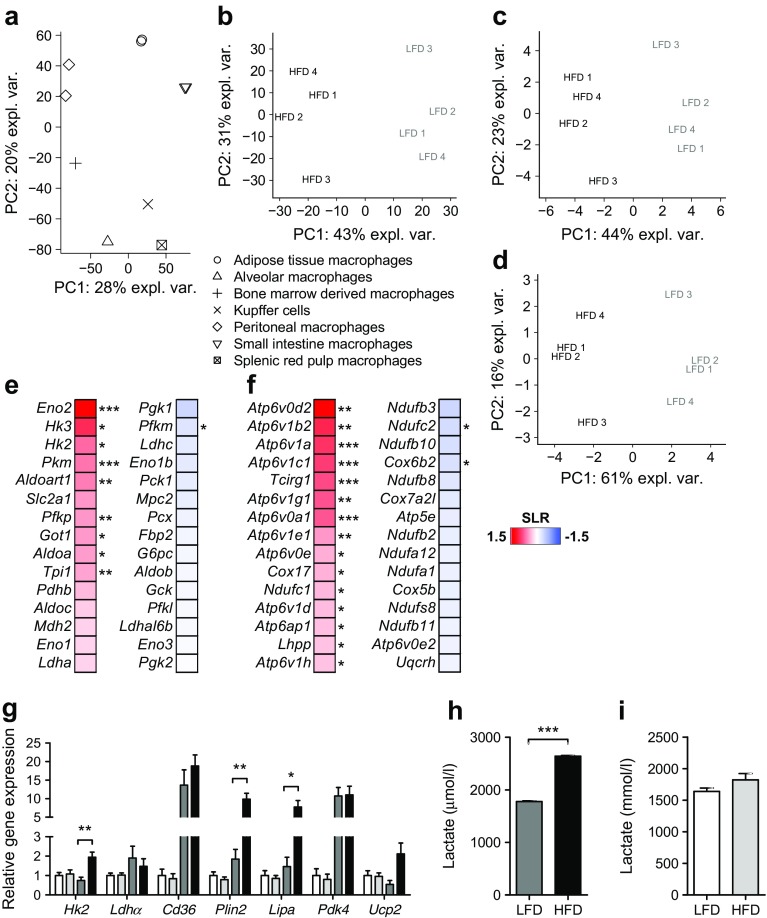
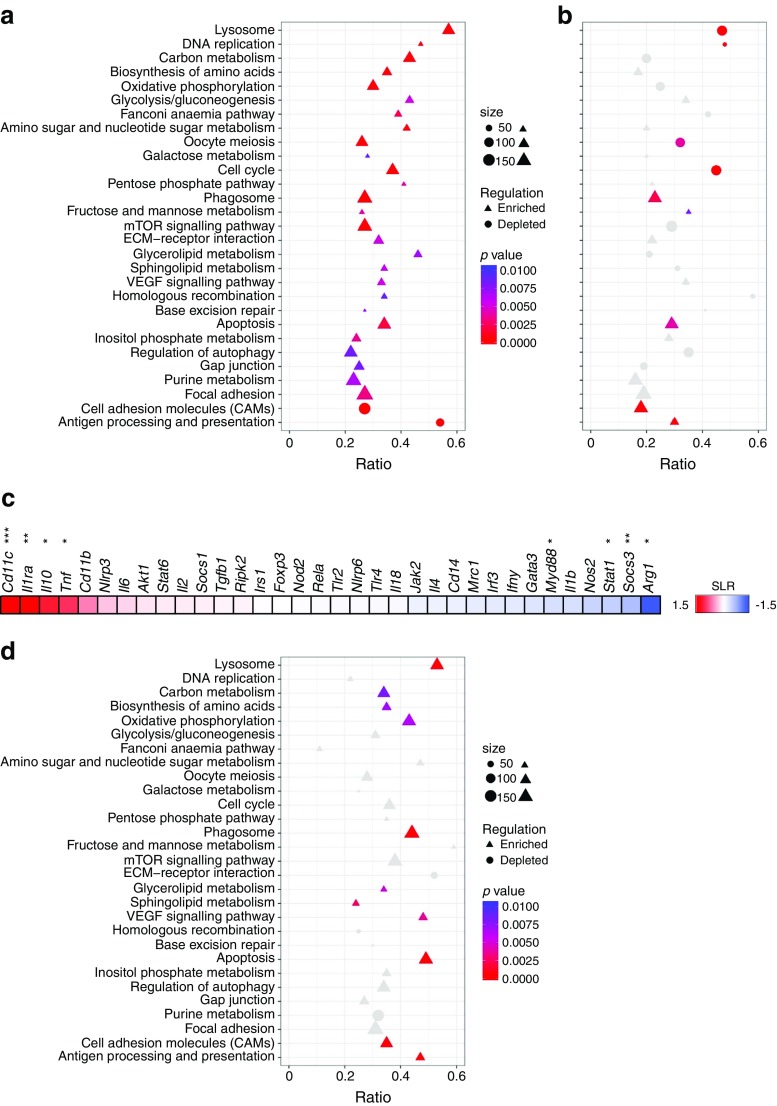
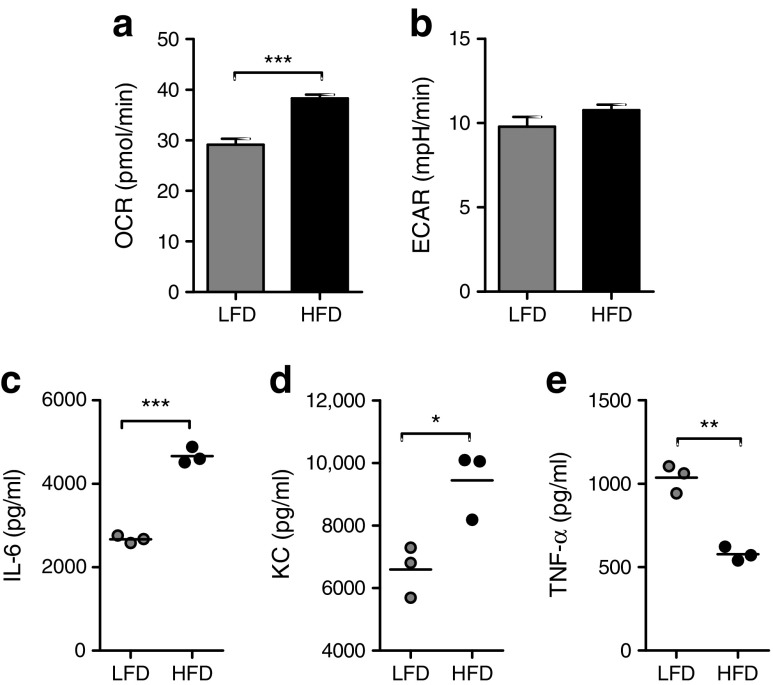
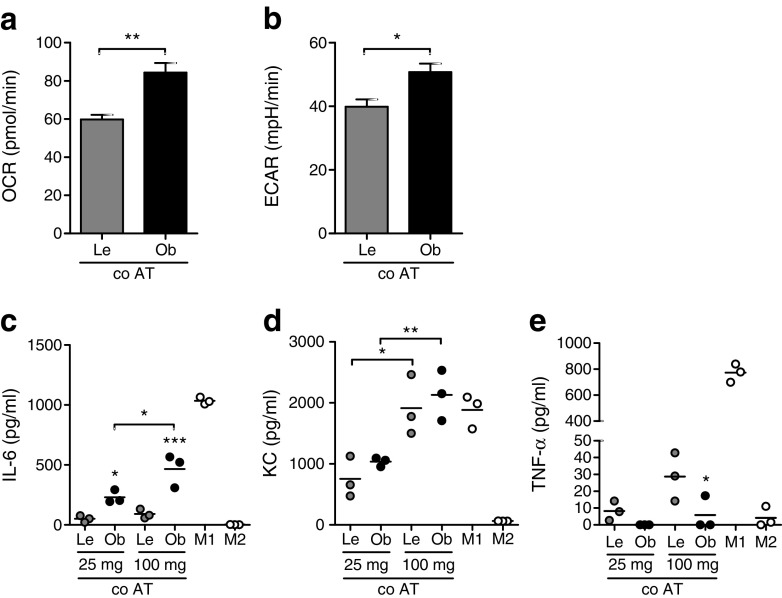
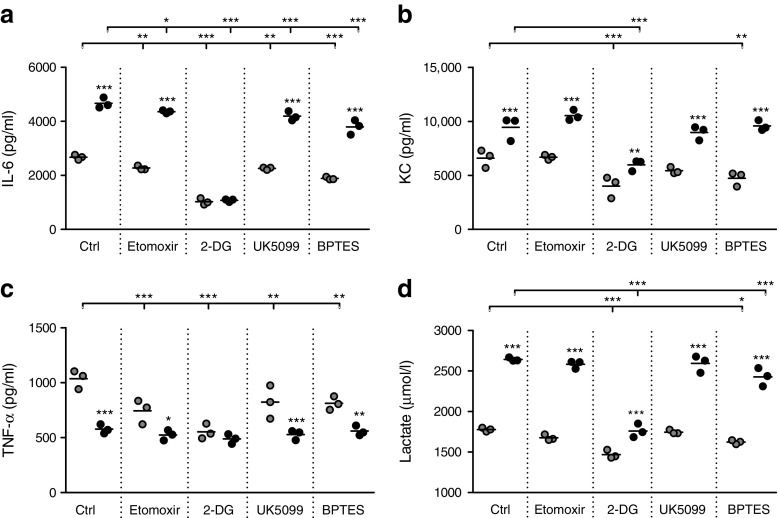
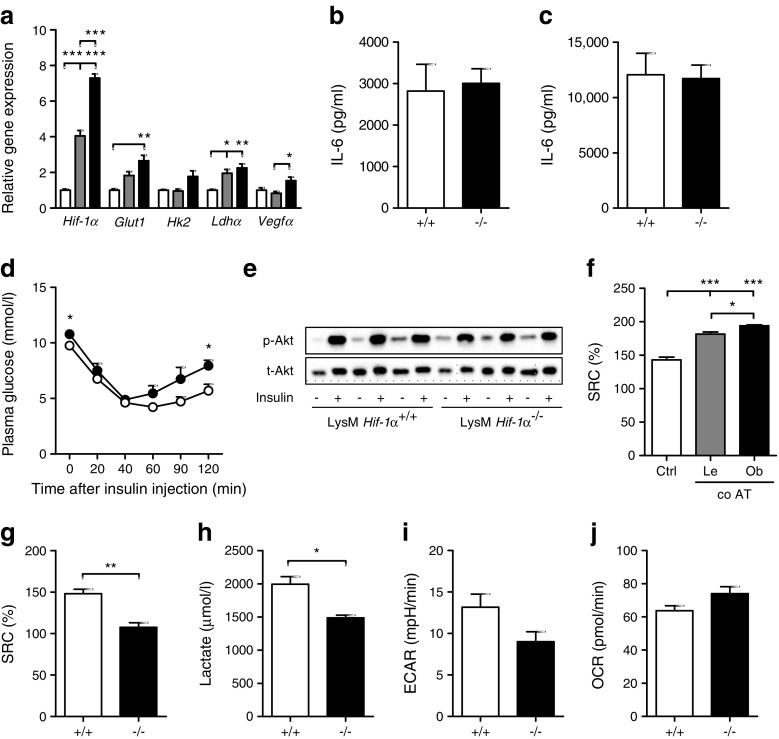
Results: Transcriptome analysis and extracellular flux measurements of mouse ATMs revealed unique metabolic rewiring in obesity characterised by both increased glycolysis and oxidative phosphorylation. Similar metabolic activation of CD14+ cells in obese individuals was associated with diabetes outcome. These changes were not observed in peritoneal macrophages from obese vs lean mice and did not resemble metabolic rewiring in M1-primed macrophages. Instead, metabolic activation of macrophages was dose-dependently induced by a set of adipose tissue-derived factors that could not be reduced to leptin or lactate. Using metabolic inhibitors, we identified various metabolic routes, including fatty acid oxidation, glycolysis and glutaminolysis, that contributed to cytokine release by ATMs in lean adipose tissue. Glycolysis appeared to be the main contributor to the proinflammatory trait of macrophages in obese adipose tissue. HIF-1α, a key regulator of glycolysis, nonetheless appeared to play no critical role in proinflammatory activation of ATMs during early stages of obesity.
Conclusions/interpretation: Our results reveal unique metabolic activation of ATMs in obesity that promotes inflammatory cytokine release. Further understanding of metabolic programming in ATMs will most likely lead to novel therapeutic targets to curtail inflammatory responses in obesity.
Data availability: Microarray data of ATMs isolated from obese or lean mice have been submitted to the Gene Expression Omnibus (accession no. GSE84000).
Keywords: Adipose tissue; Glycolysis obesity; Immunometabolism; Inflammation; Macrophages; Oxidative phosphorylation.
Conflict of interest statement
The authors declare that there is no duality of interest associated with this manuscript.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
