

The molecular basis of subtype selectivity of human kinin G-protein-coupled receptors
- PMID: 29334381
- PMCID: PMC7992120
- DOI: 10.1038/nchembio.2551
The molecular basis of subtype selectivity of human kinin G-protein-coupled receptors
Abstract
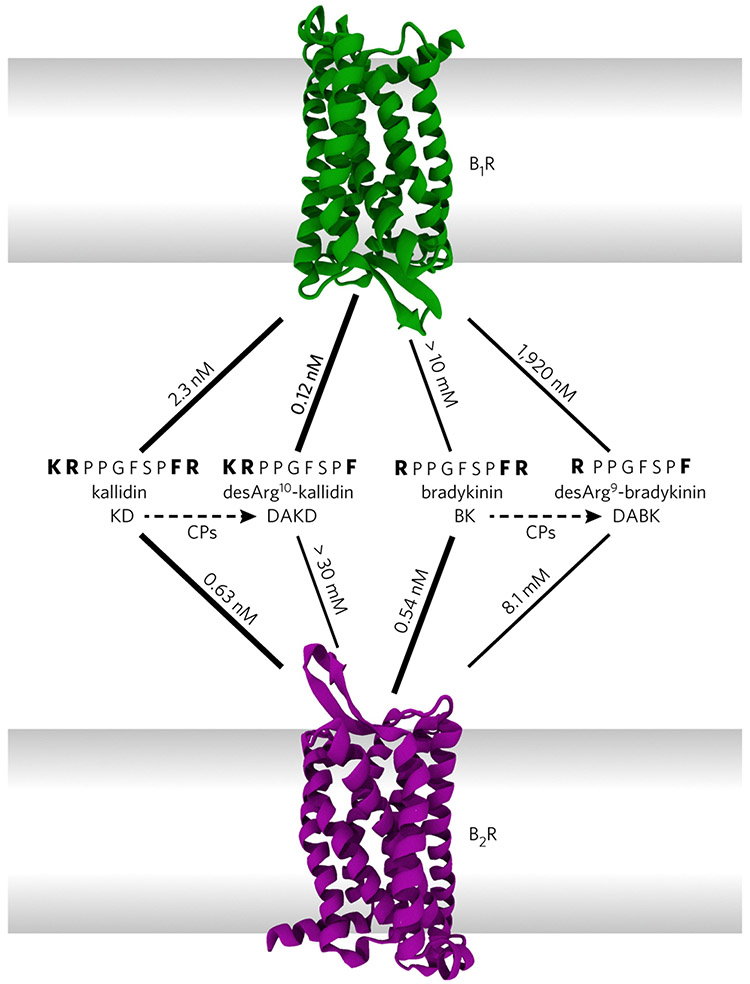
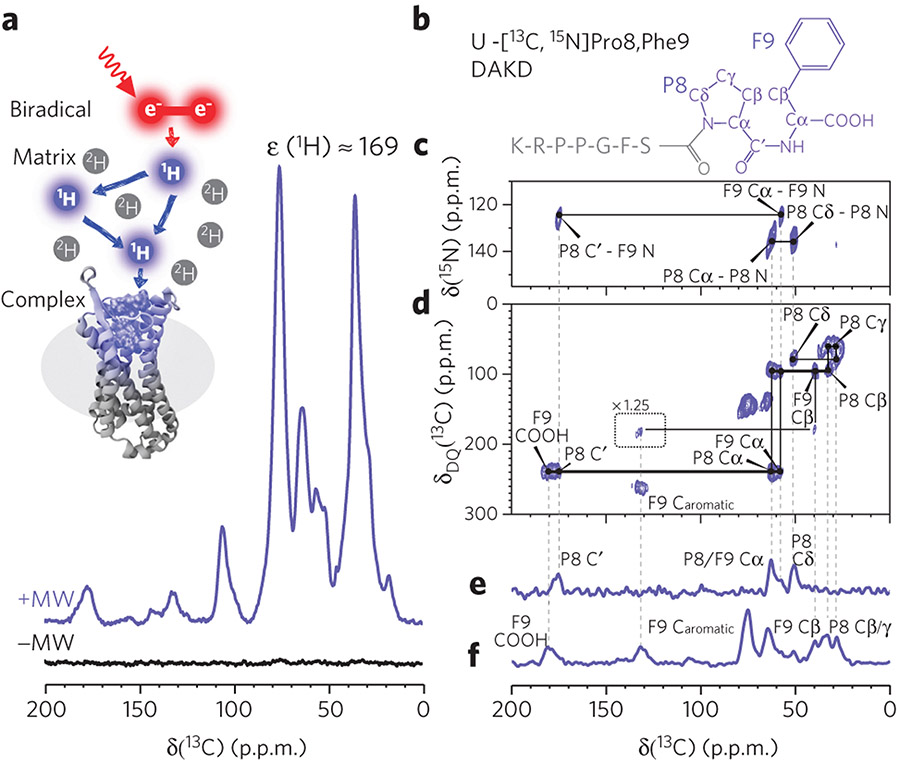
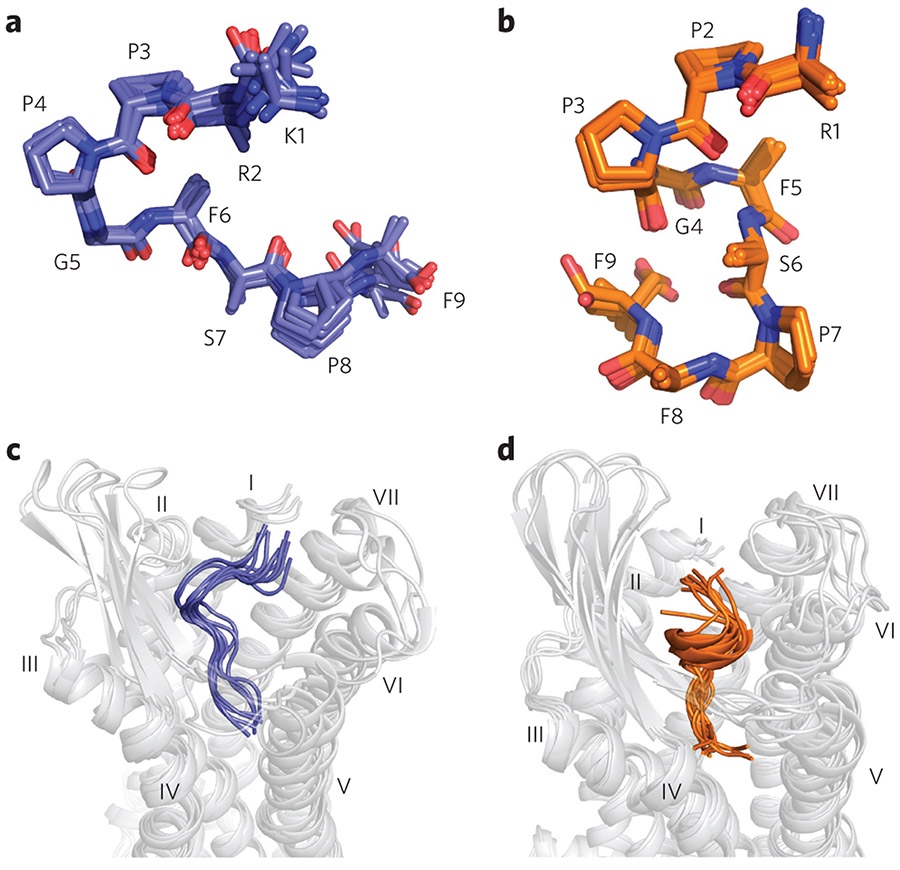
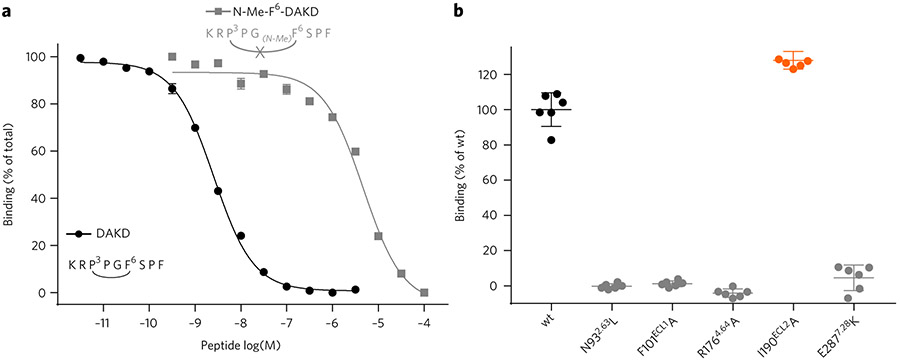
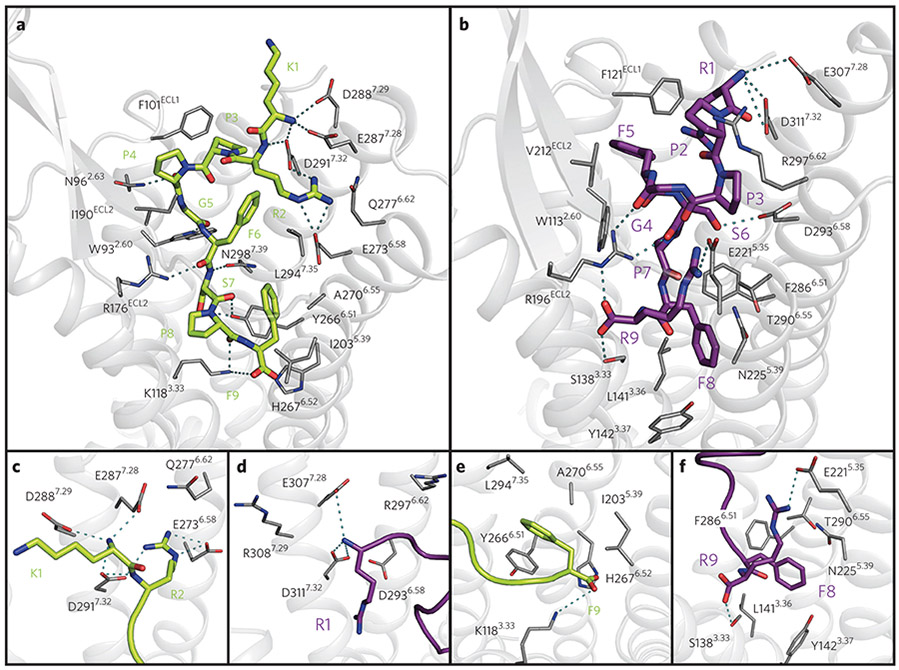
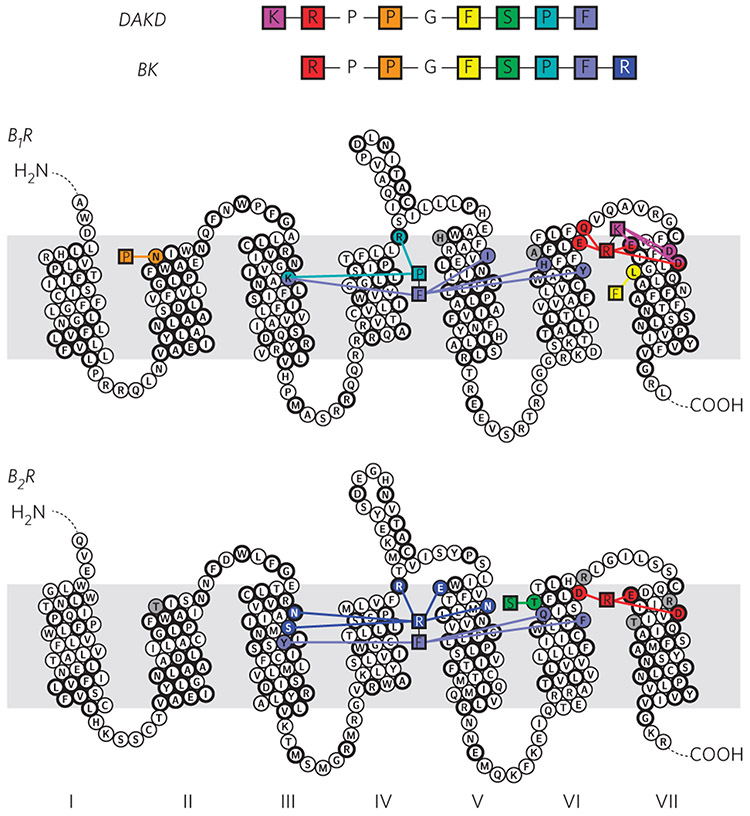
G-protein-coupled receptors (GPCRs) are the most important signal transducers in higher eukaryotes. Despite considerable progress, the molecular basis of subtype-specific ligand selectivity, especially for peptide receptors, remains unknown. Here, by integrating DNP-enhanced solid-state NMR spectroscopy with advanced molecular modeling and docking, the mechanism of the subtype selectivity of human bradykinin receptors for their peptide agonists has been resolved. The conserved middle segments of the bound peptides show distinct conformations that result in different presentations of their N and C termini toward their receptors. Analysis of the peptide-receptor interfaces reveals that the charged N-terminal residues of the peptides are mainly selected through electrostatic interactions, whereas the C-terminal segments are recognized via both conformations and interactions. The detailed molecular picture obtained by this approach opens a new gateway for exploring the complex conformational and chemical space of peptides and peptide analogs for designing GPCR subtype-selective biochemical tools and drugs.
Figures
Comment in
-
GPCR: Lock and key become flexible.Nat Chem Biol. 2018 Feb 14;14(3):201-202. doi: 10.1038/nchembio.2567. Nat Chem Biol. 2018. PMID: 29443979 No abstract available.
References
-
- Isogai S et al. Backbone NMR reveals allosteric signal transduction networks in the β1-adrenergic receptor. Nature 530, 237–241 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
