

Transcription factors orchestrate dynamic interplay between genome topology and gene regulation during cell reprogramming
- PMID: 29335546
- PMCID: PMC5810905
- DOI: 10.1038/s41588-017-0030-7
Transcription factors orchestrate dynamic interplay between genome topology and gene regulation during cell reprogramming
Abstract
Chromosomal architecture is known to influence gene expression, yet its role in controlling cell fate remains poorly understood. Reprogramming of somatic cells into pluripotent stem cells (PSCs) by the transcription factors (TFs) OCT4, SOX2, KLF4 and MYC offers an opportunity to address this question but is severely limited by the low proportion of responding cells. We have recently developed a highly efficient reprogramming protocol that synchronously converts somatic into pluripotent stem cells. Here, we used this system to integrate time-resolved changes in genome topology with gene expression, TF binding and chromatin-state dynamics. The results showed that TFs drive topological genome reorganization at multiple architectural levels, often before changes in gene expression. Removal of locus-specific topological barriers can explain why pluripotency genes are activated sequentially, instead of simultaneously, during reprogramming. Together, our results implicate genome topology as an instructive force for implementing transcriptional programs and cell fate in mammals.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
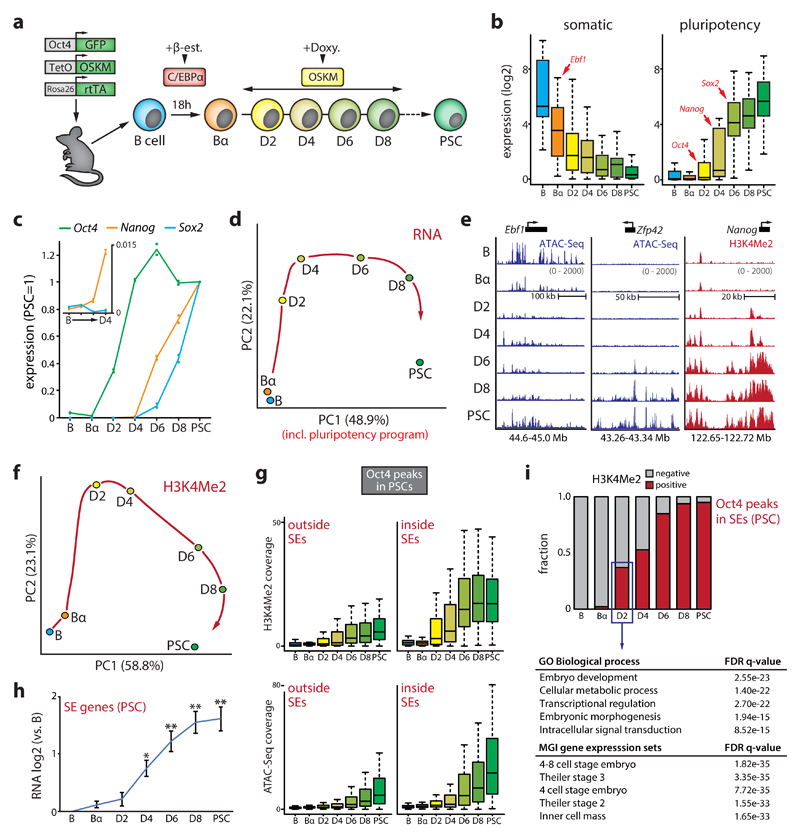
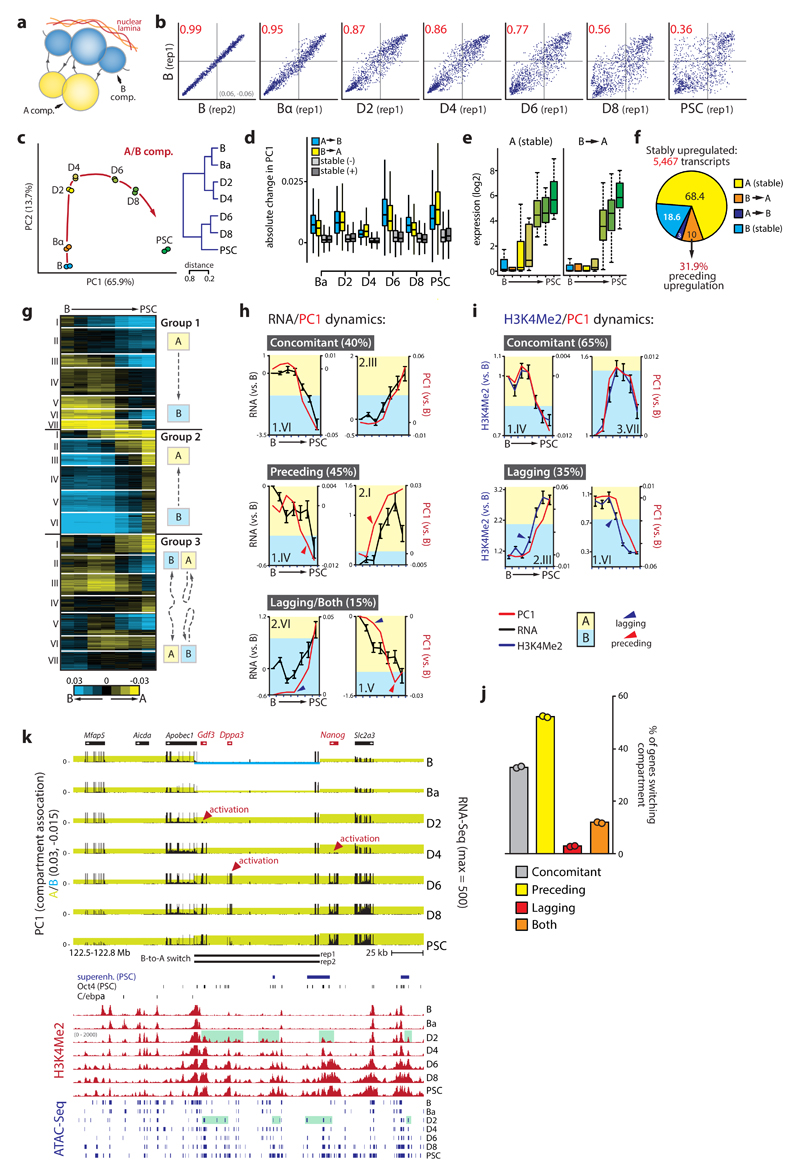
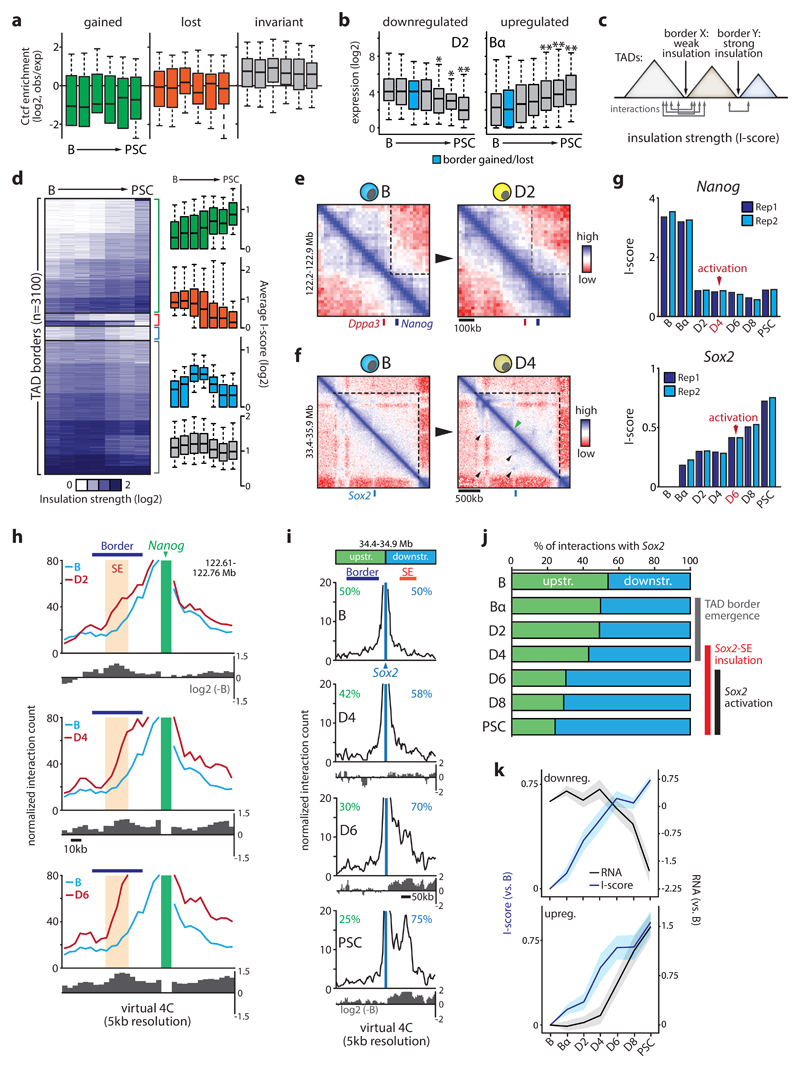
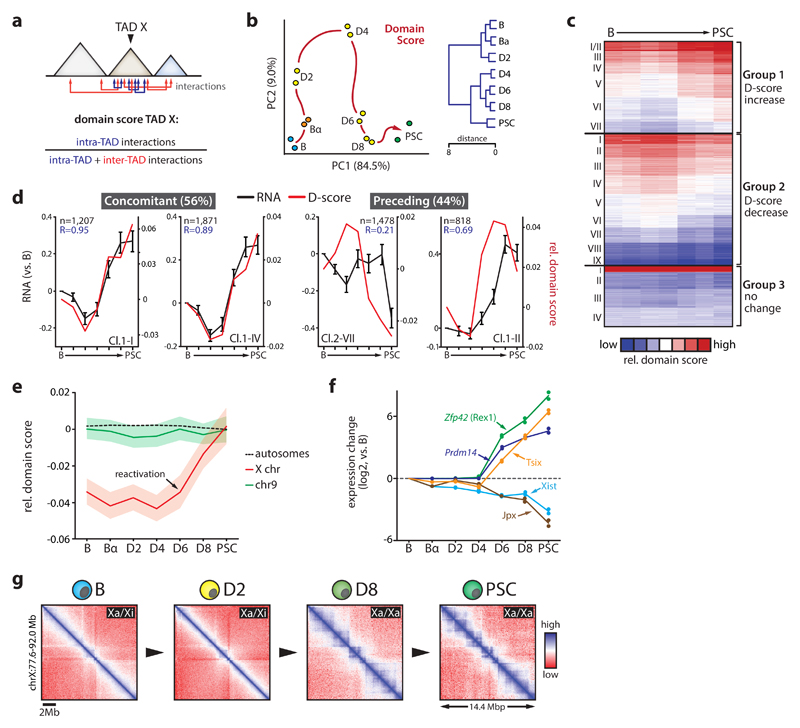
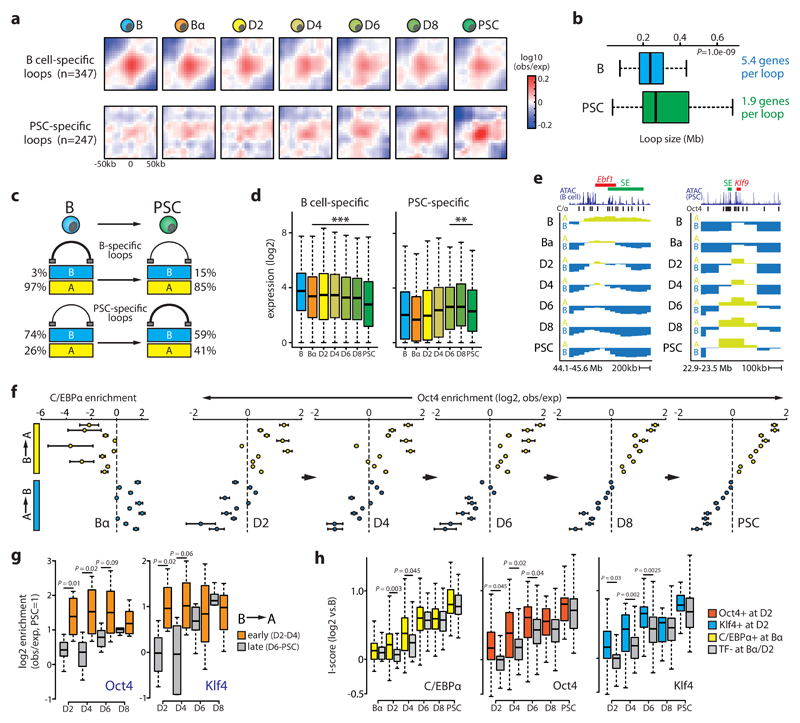
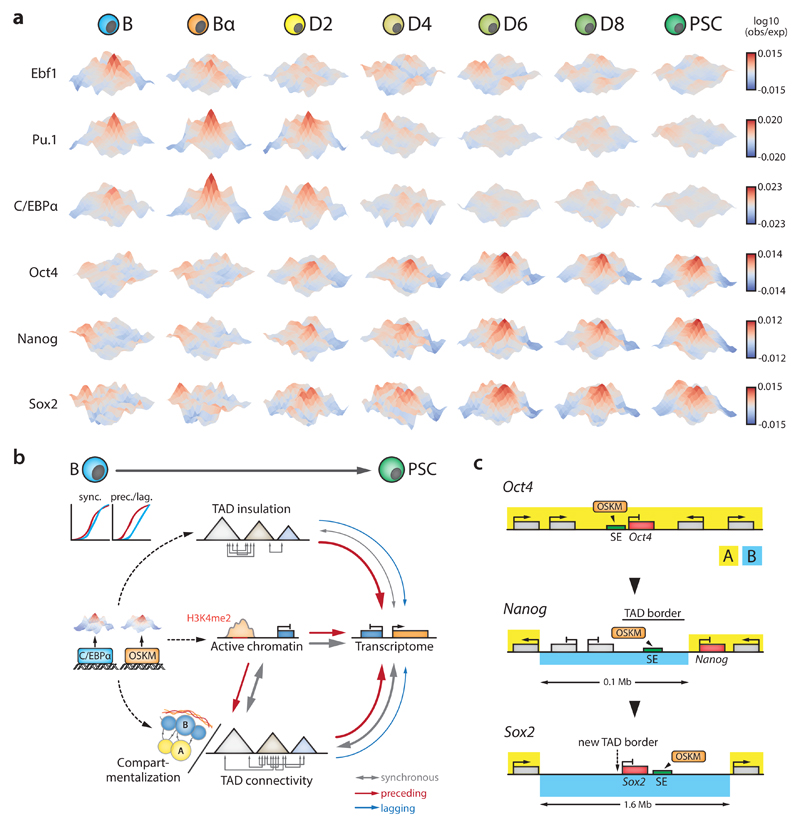
Comment in
-
Shaping the Pluripotent Genome: Switches, Borders, and Loops.Cell Stem Cell. 2018 Feb 1;22(2):148-150. doi: 10.1016/j.stem.2018.01.005. Cell Stem Cell. 2018. PMID: 29395051
References
-
- de Laat W, Duboule D. Topology of mammalian developmental enhancers and their regulatory landscapes. Nature. 2013;502:499–506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
